Neuroanatomy...Cranial nerve anatomy
•
18 recomendaciones•5,213 vistas

Neuroanatomy...Cranial nerve anatomy http://yassermetwally.com http://yassermetwally.net

Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Destacado (9)
Similar a Neuroanatomy...Cranial nerve anatomy
Similar a Neuroanatomy...Cranial nerve anatomy (20)
Más de Professor Yasser Metwally
Más de Professor Yasser Metwally (20)
Último
Último (20)
Neuroanatomy...Cranial nerve anatomy
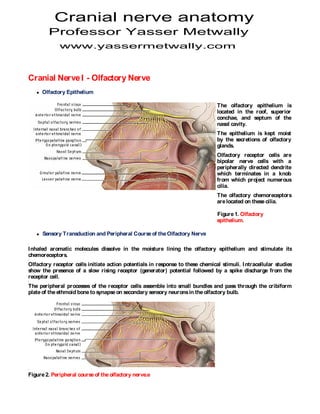
- 1. Cranial nerve anatomy Professor Yasser Metwally www.yassermetwally.com Cranial Nerve I - Olfactory Nerve Olfactory Epithelium The olfactory epithelium is located in the roof, superior conchae, and septum of the nasal cavity. The epithelium is kept moist by the secretions of olfactory glands. Olfactory receptor cells are bipolar nerve cells with a peripherally directed dendrite which terminates in a knob from which project numerous cilia. The olfactory chemoreceptors are located on these cilia. Figure 1. Olfactory epithelium. Sensory Transduction and Peripheral Course of the Olfactory Nerve I nhaled aromatic molecules dissolve in the moisture lining the olfactory epithelium and stimulate its chemoreceptors. Olfactory receptor cells initiate action potentials in response to these chemical stimuli. I ntracellular studies show the presence of a slow rising receptor (generator) potential followed by a spike discharge from the receptor cell. The peripheral processes of the receptor cells assemble into small bundles and pass through the cribiform plate of the ethmoid bone to synapse on secondary sensory neurons in the olfactory bulb. Figure 2. Peripheral course of the olfactory nerve.e
- 2. M ajor projections of the olfactory areas All three olfactory areas interface with various autonomic centers via a complex communications network. The principal pathways include: M edial forebrain bundle - provides information from all three olfactory areas to the hypothalamus. Stria medullaris- carries olfactory information from the rhinencephalon to the habenular nucleus. Stria terminalis - carries information from the amygdala to the anterior hypothalamus and preoptic area of the cortex. From the hypothalamus and habenular nucleus information is sent to the reticular formation and various cranial nerve nuclei involved in visceral responses. Examples include the dorsal motor nucleus of the vagus which mediates nausea, gastric motility and secretion, and intestinal peristalsis and the superior and inferior salivatory nuclei which mediate salivation. Figure 3. M ajor projections of the olfactory areas. Cranial Nerve I I I - Oculomotor nerve Nerve Oculomotor nerve palsy Consists of two components with distinct functions: The somatic motor component of CN I I I plays a major role in controlling the muscles responsible for the precise movement of the eyes for visual tracking or fixation on an object. The visceral motor component is involved in the pupillary light and accomodation reflexes. Supplies four of the six extraocular muscles of the Somatic motor eye and the levator (general somatic efferent) palpebrae superioris muscle of the upper eyelid. Parasympathetic Visceral motor innervation of the
- 3. (general visceral efferent) constrictor pupillae and ciliary muscles. Figure 4. Overview of occulomotor nerve components. Overview of the Somatic M otor Component There are six extraocular muscles in each orbit. The somatic motor component of CN I I I innervates the following four extraocular muscles of the eyes:
- 4. I psilateral inferior rectus muscle I psilateral inferior oblique muscle I psilateral medial rectus muscle Contralateral superior rectus muscle The remaining extraocular muscles, the superior oblique and lateral rectus muscles, are innervated by the trochlear nerve (CN I V) and abducens nerve (CN VI ), respectively. The somatic motor component of CN I I I also innervates the levator palpebrae superioris muscles bilaterally. These muscles elevate the upper eyelids. Figure 5. Overview, somatic motor component of CN I I I . Actions of the Extraocular M uscles The actions of all six extraocular muscles are summarized in the table below: Primary Secondary Tertiary M uscle I nnervation action action action Medial CN III Adduction -- -- rectus Superior CN III Elevation Intortion Adduction rectus Inferior CN III Depression Extortion Adduction rectus Inferior CN III Extorsion Elevation Abduction oblique Superior CN IV Intorsion Depression Abduction oblique Lateral CN VI Abduction -- -- rectus
- 5. A knowledge of the origins and points of insertion of the extraocular muscles on the eye relative to the axes of motion of the eye is critical to understanding the actions of these muscles. Figure 6. Origins and points of insertion of the extraocular muscles on the eye M edial Rectus M uscle The medial rectus muscle originates from the tendinous ring of the orbit and inserts on the medial border of the eye. Contraction of this muscles leads to adduction of the eye. Figure 7. M uscles of adduction. Superior Rectus M uscle The superior rectus muscle originates from the tendinous ring of the orbit and inserts on the superior surface of the eye slightly medial to the eyes vertical axis of rotation. Due to these factors, contraction of the superior rectus results in elevation, intorsion, and adduction of the eye. The primary action of the superior rectus muscle (elevation of the eye) can be isolated by having the patient look laterally and then upwards.
- 6. Figure 8. M uscles of elevation. Somatic motor component - origin and central course The somatic motor component of CN I I I originates from the oculomotor nucleus located in the rostral midbrain at the level of the superior colliculus. Like other somatic motor nuclei, the oculomotor nucleus is located near the midline just ventral to the cerebral aqueduct. I n a coronal cross-section of the brainstem the oculomotor nucleus is "V-shaped" and is bordered medially by the Edinger-Westphal nucleus and laterally and inferiorly by the medial longitudinal fasciculus which allows communication between various brainstem nuclei. Figure 9. Somatic motor component - origin and central course. Fibers leaving the occulomotor nucleus travel ventrally in the tegmentum of the midbrain passing through the red nucleus and medial portion of the cerebral peduncle to emerge in the interpeduncular fossa at the junction of the midbrain and pons.
- 7. Somatic motor component - intracranial course Upon emerging from the brainstem the oculomotor nerve passes between the posterior cerebral and superior cerebellar arteries and pierces the dura mater to enter the cavernous sinus. The nerve runs along the lateral wall of the cavernous sinus just superior to the trochlear nerve and enters the orbit via the superior orbital fissure. Figure 10. Somatic motor component - intracranial course. Somatic motor component, final innervation Within the orbit CN I I I fibers pass through the tendinous ring of the extraocular muscles and divide into superior and inferior divisions. The superior division ascends lateral to the optic nerve to innervate the superior rectus and and levator palpebrae superioris muscles on their deep surfaces. Figure 11. Somatic motor component, final innervation, anterior view. The inferior division of CN I I I splits into three branches to innervate the medial rectus and inferior rectus muscles on their ocular surfaces and the inferior oblique muscle on its posterior surface.
- 8. Figure 12. Somatic motor component, final innervation. Somatic motor component, vertical gaze The exact control of eye movements requires input from integration centers in the brain that coordinate the output from the occulomotor, trochlear, and abducens nuclei which control the six extraocular muscles. For eye movements in the vertical plane, the superior rectus, inferior rectus, inferior oblique, and superior oblique muscles of the eyes must work precisely together. The actions of these muscles is coordinated by the vertical gaze center which is thought to be located in the periaqueductal grey matter of the midbrain at the level of the superior colliculus (its location has not yet been positively identified). The vertical gaze center projects to the oculomotor nuclei which control the superior rectus, inferior rectus and inferior oblique muscles as well as to the trochlear nuclei which control the superior oblique muscles. The center that controls torsional movements of the eye is probably close to, or the same as, the vertical gaze center since all muscles that elevate or depress the eyes also cause them to rotate about their anterior-posterior axis of motion. The lateral gaze center is discussed in the abducens nerve (CN VI ) module.
- 9. Figure 13. Vertical gaze center, CN I I I & CN I V. Overview of the visceral motor component Provides parasympathetic innervation of the constrictor pupillae and ciliary muscles of the eye. The visceral motor component of CN I I I is involved in the pupillary light and accommodation reflexes. Figure 14. Overview of the visceral motor component. Visceral motor component, origin and course The visceral motor component originates from the Edinger-Westphal nucleus located in the rostral midbrain at the level of the superior colliculus. I n a coronal cross-section of the brainstem the Edinger-Westphal nucleus sit within the "V-shaped" oculomotor nuclei just ventral to the cerebral aqueduct.
- 10. Preganglionic parasympathetic fibers course ventrally through the midbrain, interpeduncular fossa, cavernous sinus, and superior orbital fissure along with the somatic motor fibers of CN I I I . Figure 15. Visceral motor component, origin and course. Visceral motor component, final innervation Once within the orbit the preganglionic parasympathetic fibers leave the nerve to the inferior oblique muscle to synapse in the ciliary ganglion which lies deep to the superior rectus muscle near the tendinous ring of the extraocular muscles. Postganglionic fibers exit the ciliary ganglion in the short ciliary nerves which enter the posterior aspect of the eye near the point of exit of the optic nerve. Within the eye these fibers travel forward between the choroid and sclera to innervate the ciliary muscles (which control the shape and therefore the refractive power of the lens) and the constrictor pupillae muscle of the iris (which constricts the pupil).
- 11. Figure 16. Visceral motor component, final innervation. Visceral motor component, pupillary light reflex Light entering the eye causes signals to be sent to the pretectal region of the midbrain via the optic nerve (CN I I ). The pretectal nucleus in turn projects bilaterally to the Edinger-Westphal nucleus. Preganglionic parasympathetic fibers from each half of the Edinger-Westphal nucleus then project to the ciliary ganglion of the ipsilateral orbit. Post-ganglionic parasympathetic fibers exit the ciliary ganglion to innervate the constrictor pupillae muscle of the ipsilateral eye. Due to the bilateral projections from the pretectal nuclei to the Edinger-Westphal nuclei, light shined into one eye produces pupillary constriction in both eyes. Direct pupillary light reflex - response in the stimulated eye. Consensual pupillary light reflex - response in the opposite eye. Visceral M otor Component, Accomodation Reflex Accommodation is an adaptation of the visual apparatus to facilitate near vision. This reflex involves the following: 1. An increase in the curvature (and therefore the refractive power) of the lens 2. Pupillary constriction to help sharpen the image on the retina 3. Convergence of the eyes to fixate on the target object The accommodation pathway is summarized below: Fibers from the primary visual cortex project, via the visual association cortex of the occipital lobe, to the superior colliculi and pretectal nuclei. Axons from the superior colliculi and pretectal nuclei project to both the Edinger-Westphal and oculomotor nuclei.
- 12. Signals from the Edinger-Westphal nuclei travel via the ipsilateral oculomotor nerve to reach the ciliary and constrictor pupillae muscles of the eye. Contraction of the ciliary muscle causes the lens to increase its curvature (and thus its refractive power), while contraction of the constrictor pupillae reduces the size of the pupillary aperture. Signals from the oculomotor nuclei travel via the ipsilateral oculomotor nerve to the medial rectus muscles causing them to contract and resulting in convergence of the eyes on the object of interest. Cranial Nerve I V - Trochlear Nerve Trochlear nerve palsy Overview The trochlear nerve has only a somatic motor component: Somatic motor innervates Somatic motor the superior oblique (general somatic efferent) muscle of the contralateral orbit. The superior oblique muscle is one of the six extraocular muscles responsible for the precise movement of the eye for visual tracking or fixation on an object. See the occulomotor nerve (CN I I I ) chapter for a discussion of eye movements and the interaction between the three nuclei and nerves that innervate the extraocular muscles.
- 13. Figure 17. Anatomic overview of the trochlear nerve. Origin and central course Figure 18. Anatomic overview of the trochlear nerve. The fibers of the trochlear nerve originate from the trochlear nucleus located in the tegmentum of the midbrain at the level of the inferior colliculus. The nucleus is located just ventral to the cerebral aqueduct. I t is readily identifiable by its close association with the myelinated medial longitudinal fasciculus that allows communication between various brainstem nuclei. Fibers leaving the trochlear nucleus travel dorsally to wrap around the cerebral aqueduct. All fibers of the two trochlear nerves decussate (i.e. cross) in the superior medullary velum and exit the dorsal surface of the brainstem just below the contralateral inferior colliculus.
- 14. Figure 19. Origin and central course. I ntracranial course Upon emerging from the dorsal surface of the brainstem the trochlear nerve curves around the brainstem in the subarachnoid space and emerges between the posterior cerebral and superior cerebellar arteries (along with CN I I I fibers). The trochlear nerve then enters and runs along the lateral wall of the cavernous sinus with CNS I I I , V, and VI . Figure 20. I ntracranial course. I ntracranial course and final innervation
- 15. From the cavernous sinus the trochlear nerve enters the orbit through the superior orbital fissure. CN I V does not pass through the tendinous ring of the extraocular muscles, rather it passes above the ring. The trochlear nerve then crosses medially along the roof of the orbit above the levator palpebrae and superior rectus muscles to innervate the superior oblique muscle along its proximal one-third: Figure 21. Final innervation of trochlear nerve. Clinical Correlation The superior oblique muscle normally depresses, intorts, and abducts the eye (fig. 4-6a). Damage to the trochlear nerve will present as: Extorsion (outward rotation) of the affected eye due to the unopposed action of the inferior oblique muscle. Vertical diplopia (double vision) due to the extorted eye. Weakness of downward gaze most noticeable on medially-directed eye. This is often reported as difficulty in descending stairs. Figure 22. Normal action of the superior oblique muscle.
- 16. Figure 23. Paralysis of the superior oblique muscle. Extortion (outward rotation) of the affected eye due to the unopposed action of the inferior oblique muscle (Fig. 4-6b). Vertical diplopia (double vision) due to the extorted eye. Weakness of downward gaze most noticeable on medially directed eye. This is often reported as difficulty in descending stairs. Head tilt (Fig. 4- 6b): patient will often tilt his head opposite the side of the affected eye in an attempt to compensate for the outwardly rotated eye. Due to its long peripheral course around the midbrain CN I V is particularly susceptible to head trauma. Cranial Nerve VI - Abducens Nerve Overview Has only a somatic motor (general somatic efferent) component. Somatic motor: innervates the lateral rectus muscle of the ipsilateral orbit. The lateral rectus muscle is one of the six extraocular muscles responsible for the precise movement of the eye for visual tracking or fixation on an object. See the CN I I I section (occulomotor nerve) for a discussion of eye movements and the interaction between the three nuclei and nerves that innervate the extraocular muscles.
- 17. Figure 24 Abducens nerve and the lateral rectus muscle. The abducens nerve innervates the lateral rectus muscle of the ipsilateral orbit. The lateral rectus muscle is responsible for lateral gaze (its contraction causes the eye to be abducted): Figure 25. Actions of the lateral rectus muscle. Origin and central course The fibers of the abducens nerve originate from the abducens nucleus located in the caudal pons at the level of the facial colliculus. The nucleus is located just ventral to the fourth ventricle near the midline. Axons of CN VI I (facial nerve) loop around the abducens nucleus and give rise to a bulge in the floor of the fourth ventricle - the facial colliculus. Fibers leaving the abducens nucleus travel ventrally to exit the brainstem at the border of the pons and medullary pyramids.
- 18. Figure 26. Origin and central course. I ntracranial course Upon exiting the brainstem the abducens nerve climbs superiorly along the ventral surface of the pons. On reaching the apex of the petrous portion of the temporal bone the nerve makes a sharp turn anteriorly to enter the cavernous sinus. The abducens nerve travels along the lateral wall of the cavernous sinus with CNS I I I , I V, and V. Figure 27. I ntracranial course of abducens nerve. I ntracranial course and final innervation From the cavernous sinus the abducens nerve enters the orbit through the superior orbital fissure. CN VI passes through the tendinous ring of the extraocular muscles and innervates the lateral rectus muscle
- 19. on its deep surface: Figure 28. Final innervation of abducens nerve. Cranial Nerve VI I - Facial Nerve Facial nerve palsy Component Overview The facial nerve has four components with distinct functions:
- 20. Supplies the muscles of facial expression; Brancial motor posterior belly of digastric (special visceral efferent) muscle; stylohyoid, and stapedius. Parasympathetic innervation of the lcrimal, submandibular, and Visceral motor sublingual glands, as well (general visceral efferent) as mucous membranes of nasopharynx, hard and soft palate. Taste sensation from the Special sensory anterior 2/3 of tongue; (special afferent) hard and soft palates. General sensation from General sensory the skin of the concha of (general somatic afferent) the auricle and from a small area behind the ear. Branchial motor fibers constitute the largest portion of the facial nerve. The remaining three components are bound in a distinct fascial sheath from the branchial motor fibers. Collectively these three components are referred to as the nervus intermedius. Component Overview
- 21. Figure 29. Functional component overview of the facial nerve. Branchial M otor Component The largest component of the facial nerve. Provides voluntary control of the muscles of facial expression (including buccinator, occipitalis and platysma muscles), as well as the posterior belly of the digastric, stylohyoid and stapedius muscles. Note the branchial motor components of the facial nerve: Figure 30. Branchial motor components of the facial nerve. Origin and Central Course
- 22. The branchial motor component originates from the motor nucleus of CN VI I in the caudal pons. Fibers leaving the motor nucleus of CN VI I initially travel medially and dorsally to loop around the ipsilateral abducens nucleus (CN VI ) producing a slight bulge in the floor of the fourth ventricle - the facial colliculus. Fibers then course so as to exit the ventrolateral aspect of the brainstem at the caudal border of the pons in conjunction with the nervus intermedius components of CN VI I . Figure 31. Brainstem section: origin and central course of the branchial motor components of the facial nerve. I ntracranial course Upon emerging from the ventrolateral aspect of the caudal border of the pons, all of the components of CN VI I enter the internal auditory meatus along with the fibers of CN VI I I (vestibulocochlear nerve). The fibers of CN VI I pass through the facial canal in the petrous portion of the temporal bone. The course of the fibers is along the roof of the vestibule of the inner ear, just posterior to the cochlea.
- 23. Figure 32. I ntracranial course- branchial motor components of the facial nerve. Facial nerve origin and inner ear anatomy. Extracranial Course and Final I nnervation The posterior auricular nerve, nerve to the posterior belly of the digastric and the nerve to the stylohyoid muscle are given off upon the facial nerve's exit from the stylomastoid foramen. The remaining fibers enter the substance of the parotid gland and divide to form the temporal, zygomatic, buccal, mandibular, and cervical branches to innervate the muscles of facial expression. Figure 33. Entracranial course- branchial motor components of the facial nerve. Geniculate ganglion and inner ear. Voluntary Control of the M uscles of Facial Expression Signals for voluntary movement of the facial muscles originate in the motor cortex (in association with other cortical areas) and pass via the corticobulbar tract in the posterior limb of the internal capsule to the motor
- 24. nuclei of CN VI I . Fibers pass to both the ipsilateral and contralateral motor nuclei of CN VI I in the caudal pons: Figure 34. Extra-cranial course and final innervation; branchial motor components of the facial nerve. Voluntary Control of the M uscles of Facial Expression The portion of the nucleus that innervates the muscles of the forehead receives corticobulbar fibers from both the contralateral and ipsilateral motor cortex. The portion of the nucleus that innervates the lower muscles of facial expression receives corticobulbar fibers from only the contralateral motor cortex. This is very important clinically as central (upper motor neuron) and peripheral (lower motor neuron) lesions will present differently.
- 25. Figure 35. Voluntary control of the muscles of facial expression. After Wilson-Pauwels, et al., 1988. Lower M otor Neuron (LM N) Lesion Results from damage to the motor nucleus of CN VI I or its axons. A LM N lesion results in the paralysis of all muscles of facial expression (including those of the forehead) ipsilateral to the lesion. Figure 36. Voluntary control of the muscles of facial expression, detail. After Wilson-Pauwels, et al., 1988. Clinical Correlation - Bell's Palsy A LM N lesion of CN VI I which occurs at or beyond the stylomastoid foramen is commonly referred to as a Bell's Palsy. Characteristic indications of a LM N lesion or Bell's Palsy include the following, on the affected side: M arked facial asymmetry Atrophy of facial muscles Eyebrow droop Smoothing out of forehead and nasolabial folds Drooping of the mouth corner Uncontrolled tearing Loss of efferent limb of conjunctival reflex (cannot close eye) Lips cannot be held tightly together or pursed Diificulty keeping food in mouth while chewing on the affected side
- 26. Figure 37. Lower M otor Neuron (LM N) Lesion. Clinical Correlation - Upper M otor Neuron (UM N) Lesion Results from damage to neuronal cell bodies in the cortex or their axons that project via the corticobulbar tract through the posterior limb of the internal capsule to the motor nucleus of CN VI I . With an UM N lesion, voluntary control of only the lower muscles of facial expression on the side contralateral to the lesion will be lost. Voluntary control of muscles of the forehead will be spared due to the bilateral innervation of the portion of the motor nucleus of CN VI I that innervates the upper muscles of facial expression. UM N lesions are usually the result of a stroke. Figure 38. Bell's Palsy: Lower M otor Neuron (LM N) Lesion. Characteristics of an UM N lesion of the facial nerve include:
- 27. Facial asymmetry Atrophy of muscles of lower portion of the face on affected side* No eyebrow droop* I ntact folds on forehead* I ntact conjunctival reflex (orbicularis oculi innervation is intact) Smoothing of nasolabial folds on affected side Lips cannot be held tightly together or pursed Difficulty keeping food in mouth while chewing on affected side * Distinguishes UM N lesions from LM N lesions. Overview of Visceral M otor Component Parasympathetic component of the facial nerve. Consists of efferent fibers which stimulate secretion from the submandibular, sublingual, and lacrimal glands, as well as the mucous membranes of the nasopharynx and hard and soft palates. Figure 39. Overview of visceral motor components of the facial nerve. Origin and Central Course The visceral motor component originates from a diffuse collection of cell bodies in the caudal pons just below the facial nucleus known as the superior salivatory nucleus. Fibers course so as to exit the ventrolateral aspect of the brainstem at the caudal border of the pons as part of the nervus intermedius portion of CN VI I . these fibers do not loop around the abducens nucleus. The nervus intermedius exits the brainstem just lateral to the branchial motor component.
- 28. Figure 40. Visceral motor component of the facial nerve: origin and central course. I ntracranial Course Upon emerging from the ventrolateral aspect of the caudal border of the pons, all of the components of CN VI I enter the internal auditory meatus along with the fibers of CN VI I I (vestibulocochlear nerve). Within the facial canal the visceral motor fibers divide into two groups to become the greater petrosal nerve and the chorda tympani: The greater petrosal nerve supplies the lacrimal, nasal, and palatine glands. The chorda tympani supplies the submandibular and sublingual glands.
- 29. Figure 41. I ntracranial course, visceral motor component of the facial nerve. Course of the Greater Petrosal Nerve At the geniculate ganglion the greater petrosal nerve turns anteriorly and medially exiting the temporal bone via the petrosal foramen and entering the middle cranial fossa. Figure 42. Course of the greater petrosal nerve through the temporal bone. The greater petrosal nerve passes deep to the trigeminal ganglion to enter the foramen lacerum. The nerve traverses the foramen and enters a canal at the base of the medial pterygoid plate in conjunction with sympathetic fibers (deep petrosal nerve) branching from the plexus following the internal carotid artery. The parasympathetic and sympathetic fibers together make up the nerve of the pterygoid canal. Upon exiting the pterygoid canal, pre-ganglionic parasympathetic fibers of CN VI I synapse in the pterygopalatine ganglion which is suspended from the fibers of the maxillary division of the trigeminal nerve (V2) in the pterygopalatine fossa. Post-ganglionic parasympathetic fibers then follow the fibers of V2 to reach the lacrimal gland (via the lacrimal nerve) and the mucous membranes of the nasal and oral pharynx.
- 30. Figure 43. Extra-cranial course of the greater petrosal nerve. Course of the Chorda Tympani The pre-ganglionic fibers of the chorda tympani branch from the other fibers of CN VI I as they pass through the facial canal just posterior to the middle ear. The fibers pass through the middle ear in close relationship with the tympanic membrane and exit the base of the skull to enter the inferotemporal fossa: Figure 44. Course of the chorda tympani, inner ear. I n the inferotemporal fossa the chorda tympani joins the fibers of the lingual branch of the mandibular division of CN V (V3). CN VI I pre-ganglionic fibers synapse in the submandibular ganglion suspended from the lingual nerve (V3). Post-ganglionic fibers then either enter the submandibular gland directly or again follow the lingual nerve before branching to innervate the sublingual gland:
- 31. Figure 45. Distal course and final innervation of the chorda tympani. Overview of Special Sensory Component Consists of afferent fibers which convey taste information from the anterior 2/3 of the tongue and the hard and soft palates. Figure 46. Facial nerve, special sensory component. Peripheral Course Chemoreceptors of the taste buds located on the anterior 2/3 of the tongue and hard and soft palates initiate receptor (generator) potentials in response to chemical stimuli. The taste buds synapse with the peripheral processes of special sensory neurons from CN VI I . These neurons generate action potentials in response to the taste bud's receptor potentials. The peripheral processes of these neurons follow the lingual nerve and then chorda tympani to the petrous portion of the temporal bone (similar to the path followed by the efferent visceral motor fibers).
- 32. Figure 47. Chorda tympani and the lingual nerve. The cell bodies of these primary afferent neurons reside in the geniculate ganglion: Figure 48. Geniculate ganglion.