
Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Similar a La Célula eucariota
Similar a La Célula eucariota (20)
Más de Colegio Salesiano Sto. Domingo Savio. Monzón. Huesca. Spain
Más de Colegio Salesiano Sto. Domingo Savio. Monzón. Huesca. Spain (18)
Último
Último (20)
La Célula eucariota
- 1. Antonio Campo Buetas Biología 2º Bachillerato
- 2. ÍNDICE • Esquema general de la célula eucariota. Diferencias entre célula eucariota vegetal y animal. La pared celular. • La membrana celular o plasmática. Modelo de mosaico fluido o de Singer-Nicholson. • El transporte a través de la membrana: transporte activo y pasivo (difusión simple y difusión facilitada). • Dinámica de la membrana: fagocitosis, endocitosis y exocitosis. • El citoplasma: hialoplasma (o citosol) y orgánulos citoplasmáticos. • Orgánulos citoplasmáticos: retículo endoplasmático, ribosomas, aparato de Golgi, lisosomas, mitocondrias, cloroplastos, vacuolas, peroxisomas. • El núcleo: la envoltura nuclear, el nucleoplasma, nucleolos, cromatina/cromosomas. • El citoesqueleto. Microtúbulos; centrosoma, cilios y flagelos. Función de los microfilamentos (idea general).
- 3. LA TEORÍA ENDOSIMBIÓTICA • La teoría endosimbiótica de Whittaker y Margulis postula que la célula eucariota evolucionó a partir de una célula procariota emparentada con las arqueobacterias. • Este proceso debió ocurrir según el registro fósil hace entre 2000-1500 ma. • Ese procariota ancestral a través de procesos de invaginación de la membrana plasmática creo un sistema de endomembranas que le sirvió, también, para proteger su información genética con una membrana nuclear. • Más adelante, incorporó mediante endocitosis a bacterias aerobias con las que estableció una relación simbiótica que le permitieron ser más eficiente en la obtención de energía utilizando el oxígeno en la respiración celular. Estas bacterias aerobias se transformarán con el tiempo en las actuales mitocondrias. • Posteriormente, incorporó por endocitosis bacterias fotosintéticas que le permitieron obtener materia orgánica a partir de materia inorgánica aprovechando la energía de la luz. Estas bacterias fotosintéticas a la larga se convirtieron en los cloroplastos actuales. • Recordamos que la simbiosis es una relación interespecífica, entre dos especies diferentes, en las que ambas salen beneficiadas. La célula hospedadora obtuvo mayor energía con las bacterias aerobias y/o la posibilidad de utilizar la energía de la luz para sintetizar materia orgánica a partir de materia inorgánica, mientras que estas bacterias encontraban un ambiente de protección con todos los nutrientes que necesitaban para realizar sus procesos vitales de nutrición y reproducción.
- 4. PRUEBAS QUE APOYAN LA TEORÍA ENDOSIMBIÓTICA • Las mitocondrias y los cloroplastos son orgánulos que presentan doble membrana. La membrana externa correspondería a la membrana del proceso de entrada en la célula ancestral por endocitosis. • Mitocondrias y cloroplastos conservan un cromosoma circular como el de las bacterias. • Mitocondrias y cloroplastos tienen ribosomas 70 S como los de las bacterias. • Estos orgánulos son capaces de dividirse dentro de las células eucariotas por división binaria como las bacterias. • En la actualidad conocemos ejemplos de protozoos, organismos unicelulares, que incorporan a su interior bacterias aerobias o bacterias fotosintéticas con las que mantienen una relación simbiótica. Por lo tanto se trata de un proceso y una relación que bien pudo darse en los orígenes de la célula eucariota.
- 7. • Las células vegetales suelen ser más grandes, tienen pared celular que les da un aspecto poligonal, una gran vacuola central y cloroplastos. No tienen centriolos. • La célula eucariota animal presenta centriolos, carece de pared celular y de cloroplastos y suele tener vacuolas pequeñas.
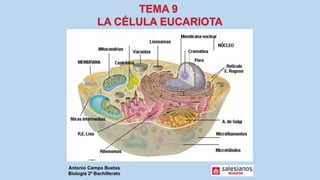
- 19. Célula eucariota animal con sus orgánulos y las funciones que desempeñan
- 27. Las células vegetales poseen una envuelta externa a la membrana plasmática, altamente organizada y rígida, que constituye la pared celular y cumple las siguientes funciones: • Confiere rigidez al vegetal y contribuye al mantenimiento de la forma celular. • Une las células adyacentes, conectando las células de los tejidos vegetales. • Posibilita el intercambio de fluidos y la comunicación intercelular. • Permite a las células vegetales vivir en el medio hipotónico de la planta, impidiendo que éstas se hinchen y lleguen a estallar. • Impermeabiliza la superficie vegetal en algunos tejidos gracias a la cutina y la suberina para evitar las pérdidas de agua. • Sirve de barrera al paso de agentes patógenos.
- 28. ESTRUCTURA Y COMPOSICIÓN DE LA PARED CELULAR: La pared celular nos recuerda al encofrado de una edificación. Tiene dos componentes principales: Un componente cristalino fibrilar, constituido por fibras de celulosa (los cables de acero en la construcción); y un componente amorfo, formado por una matriz de pectinas, hemicelulosa, agua, sales minerales y, en algunos casos, ligninas (el hormigón de la construcción). En las células diferenciadas, la pared celular aparece como una estructura gruesa compuesta por varias capas que se van depositando a medida que madura la célula. Estas capas son: • Lámina media • Pared primaria • Pared secundaria
- 29. ESTRUCTURA Y COMPOSICIÓN: • Lámina media: Es la capa más externa y la primera que se forma después de la división celular, y puede ser compartida por las células adyacentes de un tejido. Está integrada por pectinas y proteínas, que se unen posteriormente a iones Ca2+. • Pared primaria: Se trata de una gruesa capa de estructura fibrilar, situada por debajo de la lámina media hacia el interior de la célula. Está constituida, fundamentalmente, por largas fibras de celulosa cohesionadas por polisacáridos (hemicelulosa y pectinas) y glucoproteínas. • Pared secundaria: El la capa más interna y se encuentra por debajo de la pared primaria en algunos tipos especiales de células vegetales ( tejidos de soporte o vasculares ). Consta de una o varias capas fibrilares, semejantes en composición a la pared primaria, aunque contienen celulosa en mayor proporción y carecen de pectinas. En estas capas fibrilares, las miofibrillas de celulosa se ordenan paralelamente dando lugar a varias capas con diferente ordenación.
- 30. ESTRUCTURA Y COMPOSICIÓN: • Lámina media: Es la capa más externa y la primera que se forma después de la división celular, y puede ser compartida por las células adyacentes de un tejido. Está integrada por pectinas y proteínas, que se unen posteriormente a iones Ca2+. • Pared primaria: Se trata de una gruesa capa de estructura fibrilar, situada por debajo de la lámina media hacia el interior de la célula. Está constituida, fundamentalmente, por largas fibras de celulosa cohesionadas por polisacáridos (hemicelulosa y pectinas) y glucoproteínas. • Pared secundaria: El la capa más interna y se encuentra por debajo de la pared primaria en algunos tipos especiales de células vegetales ( tejidos de soporte o vasculares ). Consta de una o varias capas fibrilares, semejantes en composición a la pared primaria, aunque contienen celulosa en mayor proporción y carecen de pectinas. En estas capas fibrilares, las miofibrillas de celulosa se ordenan paralelamente dando lugar a varias capas con diferente ordenación.
- 31. ESTRUCTURA Y COMPOSICIÓN: • Lámina media: Es la capa más externa y la primera que se forma después de la división celular, y puede ser compartida por las células adyacentes de un tejido. Está integrada por pectinas y proteínas, que se unen posteriormente a iones Ca2+. • Pared primaria: Se trata de una gruesa capa de estructura fibrilar, situada por debajo de la lámina media hacia el interior de la célula. Está constituida, fundamentalmente, por largas fibras de celulosa cohesionadas por polisacáridos (hemicelulosa y pectinas) y glucoproteínas. • Pared secundaria: El la capa más interna y se encuentra por debajo de la pared primaria en algunos tipos especiales de células vegetales ( tejidos de soporte o vasculares ). Consta de una o varias capas fibrilares, semejantes en composición a la pared primaria, aunque contienen celulosa en mayor proporción y carecen de pectinas. En estas capas fibrilares, las miofibrillas de celulosa se ordenan paralelamente dando lugar a varias capas con diferente ordenación.
- 32. ESTRUCTURA Y COMPOSICIÓN: • Lámina media: Es la capa más externa y la primera que se forma después de la división celular, y puede ser compartida por las células adyacentes de un tejido. Está integrada por pectinas y proteínas, que se unen posteriormente a iones Ca2+. • Pared primaria: Se trata de una gruesa capa de estructura fibrilar, situada por debajo de la lámina media hacia el interior de la célula. Está constituida, fundamentalmente, por largas fibras de celulosa cohesionadas por polisacáridos (hemicelulosa y pectinas) y glucoproteínas. • Pared secundaria: El la capa más interna y se encuentra por debajo de la pared primaria en algunos tipos especiales de células vegetales ( tejidos de soporte o vasculares ). Consta de una o varias capas fibrilares, semejantes en composición a la pared primaria, aunque contienen celulosa en mayor proporción y carecen de pectinas. En estas capas fibrilares, las miofibrillas de celulosa se ordenan paralelamente dando lugar a varias capas con diferente ordenación.
- 33. ESTRUCTURA Y COMPOSICIÓN: • Lámina media: Es la capa más externa y la primera que se forma después de la división celular, y puede ser compartida por las células adyacentes de un tejido. Está integrada por pectinas y proteínas, que se unen posteriormente a iones Ca2+. • Pared primaria: Se trata de una gruesa capa de estructura fibrilar, situada por debajo de la lámina media hacia el interior de la célula. Está constituida, fundamentalmente, por largas fibras de celulosa cohesionadas por polisacáridos (hemicelulosa y pectinas) y glucoproteínas. • Pared secundaria: El la capa más interna y se encuentra por debajo de la pared primaria en algunos tipos especiales de células vegetales ( tejidos de soporte o vasculares ). Consta de una o varias capas fibrilares, semejantes en composición a la pared primaria, aunque contienen celulosa en mayor proporción y carecen de pectinas. En estas capas fibrilares, las miofibrillas de celulosa se ordenan paralelamente dando lugar a varias capas con diferente ordenación.
- 34. En ocasiones, entran a formar parte de su composición polímeros, como la lignina (xilema y esclerénquima), ceras y cutina (haz de las hojas) o suberina (corcho). En algunas paredes se observan también inclusiones minerales, principalmente carbonatos y sílice. • Lignificación: impregnación de la pared con lignina, que le proporciona mayor rigidez sin disminución de su permeabilidad. Suele darse en los tejidos de sostén y en algunos tejidos conductores, colaborando en mantener erguido el cuerpo de la planta. • Cutinización y suberificación: impregnación por cutina o por suberina respectivamente, sustancia que confieren impermeabilidad a la pared en los tejidos protectores. La cutinización se presenta en la superficie externa de las células epidérmicas, mientras que las células suberificadas constituyen el tejido suberoso o corcho. Xilema (lignina) corcho (suberina) Composición del xilema y esclerénquima Cutícula de la epidermis (cutina)
- 35. ESPECIALIZACIONES EN LA PARED CELULAR Aunque la lámina media y la pared primaria son permeables al agua y a las pequeñas moléculas disueltas en ella, el paso de sustancias está facilitado por la existencia de ciertas especializaciones, como las punteaduras y los plasmodesmos.
- 36. PUNTEADURAS Y PLASMODESMOS: • Las punteaduras se pueden definir como interrupciones de la pared secundaria y, en los vegetales pluricelulares, normalmente las punteaduras de células contiguas se corresponden. Así, a nivel de las punteaduras, las células vecinas están separadas por la lámina media y las paredes primarias depositadas a ambos lados; este conjunto se llama membrana oclusiva. • Plasmodesmos: Las membranas oclusivas están perforadas como una criba y atravesadas por finísimos conductos citoplasmáticos o plasmodesmos, donde la membrana plasmática de cada célula se continúa con la de su vecina. Los plasmodesmos permiten el paso de pequeñas moléculas entre células adyacentes y posibilitan que una planta no sea un conjunto de células individualizadas, sino una unidad fisiológica. Punteaduras Membrana Oclusiva Membrana Oclusiva con Plasmodesmos
- 38. Los plasmosdesmos son esenciales para transportar moléculas entre las células de las plantas y también desde y hacia los haces vasculares. Moléculas como agua, azúcares, hormonas, proteínas y ARNs. Este tipo de viaje de citoplasma en citoplasma se denomina vía simplasto y mantiene conectadas a las células vegetales. Este transporte se puede regular disminuyendo la luz de los plasmodesmos mediante la acumulación o eliminación de calosa. Este proceso es rápido pues ocurre en unos pocos minutos Cuando una célula de una planta sufre una infección se cierran sus plasmodesmos para evitar que esta se transmita a las células vecinas.
- 39. LA MEMBRANA PLASMÁTICA • La membrana plasmática es una estructura que rodea y delimita completamente a la célula y constituye una "barrera" selectiva que controla el intercambio de sustancias entre el interior celular y el exterior. • En cortes ultrafinos, visualizados con el Microscopio electrónico de transmisión, aparece como dos bandas oscuras separadas por una banda clara, con un espesor de 7,5 nm. • Posee la misma estructura en todas las células. Esta organización es común, además, al resto de las membranas biológicas constituyentes o limitantes de los orgánulos celulares, por lo que se denomina unidad de membrana.
- 40. LA MEMBRANA PLASMÁTICA • La estructura trilaminar observada se corresponde con una bicapa lipídica con proteínas embebidas en ella. • Los lípidos se disponen en una bicapa con las zonas hidrófilas (grupos polares) hacia fuera, mientras que las zonas hidrófobas quedan enfrentadas hacia el interior. • Las membranas presentan, por tanto, dos caras: una cara externa y una cara interna que, en el caso de la membrana plasmática, está en contacto con el citoplasma celular. • Las proteínas pueden ser periféricas y estar asociadas a la cara interna o externa, o ser integrales (atraviesan la membrana totalmente).
- 41. COMPOSICIÓN DE LA MEMBRANA PLASMÁTICA La membrana plasmática que delimita a la célula eucariota está constituida por lípidos y proteínas en una proporción, por lo general, en torno al 50%. Lípidos de membrana: • Fosfolípidos: Son los lípidos más abundantes en las membranas biológicas. Son moléculas anfipáticas, presentan una zona hidrófila (cabeza polar), y una zona hidrófoba (cola apolar). • Glucolípidos: Semejantes a los fosfolípidos, pero contienen oligosacáridos. En las células animales suelen ser derivados de esfingolípidos. En las células vegetales derivan de los fosfoglicéridos. Sólo aparecen en la cara externa de la membrana plasmática, contribuyendo, en parte, a constituir el glicocaliz. • Esteroles: Derivados del colesterol y presentes en la membrana plasmática de las células eucariotas, son más abundantes, por lo general, en las células animales. Proteínas de membrana: Desempeñan funciones estructurales, de reconocimiento, transporte o metabólico. Pueden ser: • Integrales: incluidas en la membrana de lado a lado y pueden ser glucoproteínas con una parte glucídica. • Periféricas: Son proteínas unidas débilmente a la membrana, siendo más frecuentes en la parte interna. La parte interna de la membrana suele estar asociada a diferentes tipos de filamentos del citoesqueleto.
- 44. ESTRUCTURA ASIMÉTRICA DE LA MEMBRANA La cara citoplasmática y la cara externa presentan algunas diferencias en su composición química. • Por la cara externa predominan los fosfolípidos fosfatidilcolina y esfingomielina, mientras que por la cara interna los más abundantes son la fosfatidilserina y la fosfatidiletanolamina. • En la cara citoplasmática predomina una carga electroquímica negativa, por la abundancia de fosfatidilserina. • Los ácidos grasos de los fosfolípidos de la cara citoplasmática presentan más dobles enlaces y, por lo tanto, esta cara es más fluida. • Sólo se encuentran glucolípidos en la cara externa de la bicapa (Glucocáliz) más frecuente en células animales. Glucocáliz
- 45. b) MODELO DEL MOSAICO FLUIDO • El Modelo del Mosaico Fluido se debe a Singer y Nicholson en 1972. Estos autores señalaron que la membrana está formada por un mosaico de teselas (lípidos y proteínas) que constituye una estructura fluida, puesto que permite el movimiento de los lípidos y las proteínas dentro de la bicapa lipídica. • Al igual que los lípidos, las proteínas integrales poseen una naturaleza anfipática: presentan zonas hidrófilas e hidrófobas, por lo que pueden estar embebidas en la bicapa de forma integral o periférica. • En la bicapa lípidica, estas moléculas pueden presentar los siguientes movimientos: difusión lateral, rotación y flexión de las cadenas hidrocarbonadas. Es poco frecuente el movimiento en flip-flop. • Por su parte, las proteínas presentan difusión lateral a lo largo de la membrana y rotación alrededor de su eje. • La fluidez de la membrana tiene una gran importancia en la regulación del transporte entre el interior y el exterior. FUNCIONES DE LA MEMBRANA PLASMÁTICA La membrana plasmática delimita a la célula, le da forma, en el caso de las células animales, pero sobre todo regula el intercambio de sustancias entre el exterior y el interior.
- 46. MODELO DEL MOSAICO FLUIDO La fluidez de la membrana tiene una gran importancia con relación a la actividad funcional de las membranas biológicas. La mayor o menor fluidez de la membrana depende de varios factores: • Aumenta con la presencia de ácidos grasos insaturados. • Aumenta cuanto más cortas son las cadenas hidrocarbonadas de los ácidos grasos. • Aumenta con la temperatura. • Disminuye con la presencia de colesterol: El colesterol es una molécula plana y rígida, por lo que estabiliza las membranas, es decir, las hace menos flexibles y fluidas. Esta molécula se encuentra prácticamente en todas las membranas plasmáticas de las células eucariotas, siendo más abundante, en las animales.
- 47. ALGUNAS FUNCIONES DE LAS PROTEÍNAS DE MEMBRANA Las proteínas de la membrana desempeñan diversas funciones: de anclaje, de transporte (pasivo y activo), enzimática, de transducción de señales, de reconocimiento de células y de unión entre células.
- 51. TIPOS DE TRANSPORTE La principal función de la membrana plasmática es regular el transporte, es decir, el intercambio de sustancias entre el exterior y el interior de la célula. Las membranas celulares son selectivamente permeables o semipermeables y regulan el transporte celular, pues permiten el paso de determinadas moléculas o iones y restringen el de otros: • Algunas moléculas no polares de pequeño tamaño (como el oxigeno y nitrógeno molecular), moléculas polares sin carga pequeñas (como el agua o el dióxido de carbono) o solubles en lípidos (ácidos grasos y alcoholes) pueden atravesar la membrana libremente. • Las moléculas con carga, como los ácidos orgánicos, aminoácidos y otros iones (H+, Na+, Cl- K+, etc), no pueden atravesar la membrana y tienen que utilizar proteínas de transporte específicas. Podemos distinguir tres tipos básicos de transporte: • Transporte pasivo (se realiza a favor de gradiente y no requiere gasto de energía) • Transporte activo (se realiza en contra de gradiente y requiere gasto de energía) • Transporte en masa (endocitosis y exocitosis) que también requiere energía.
- 53. TIPOS DE TRANSPORTE • La difusión simple se realiza a favor de gradiente, sin gasto de energía y no requiere transportador. La realizan moléculas apolares (lípidos) y polares pequeñas sin carga (H2O) y los gases (O2, CO2…) • La difusión facilitada se realiza a favor de gradiente, sin gasto de energía y requiere la presencia de una proteína transportadora (de canal o con cambio de conformación al unirse al sustrato) Son transportados así los iones con carga, los monosacáridos, aminoácidos… • El transporte activo se realiza en contra de un gradiente, con gasto de energía y la presencia de una proteína transportadora que actúa como bomba (bomba Na-K en el transporte del impulso nervioso en las neuronas).
- 54. La ósmosis es una forma de transporte por difusión simple del agua a lo largo de una membrana plasmática que separa dos concentraciones de sales distintas. La membrana es semipermeable, es decir, deja pasar las moléculas de agua pero es impermeable a los iones con carga. Isotonía: las concentraciones de sales a ambos lados son iguales y el agua entra y sale en equilibrio. Turgencia: el medio externo presenta menor concentración de sales (hipotónico o hipoosmótico) que el medio interno (hipertónico o hiperosmótico). El agua entra hinchando a la célula animal con el riesgo de rotura de la membrana (lisis celular) o hinchando la gran vacuola central (células vegetales). Plasmólisis: el medio externo presenta mayor concentración de sales (medio hiperosmótico) que el medio interno. El agua sale de la célula y se arruga la membrana plasmática (o la membrana de la vacuola en células vegetales). Fenómenos osmóticos en glóbulos rojos
- 57. TIPOS DE TRANSPORTE POR DIFUSIÓN FACILITADA Se efectúa mediante proteínas transportadoras a favor de gradiente sin gasto de energía. Distinguimos lo siguientes tipos: • Uniporte: Una única sustancia en un sentido determinado. • Dos sustancias distintas: • Simporte: en el mismo sentido. • Antiporte: en sentido opuesto. Las proteínas transportadoras presentan una alta especificidad para el transporte de un determinado tipo de compuesto químico. En muchos casos, el transporte se lleva a cabo mediante un cambio en la conformación de la proteína. Estas proteínas suelen ser transmembranales. El cambio de conformación permite la transposición de los sitios de unión al ligando desde el exterior (estado pong) al interior de la célula (estado ping).
- 58. TRANSPORTE ACTIVO Se realiza en contra de un gradiente de concentración o presión e implica consumo de energía, proporcionada por el ATP y un transportador. El mecanismo sería similar al de la difusión facilitada, pero el cambio de conformación va ligado a la hidrólisis de ATP, por lo que las proteínas de transporte se denominan bombas: impulsan el transporte de solutos en contra de gradiente electroquímico. Un buen ejemplo es la bomba de Na+ / K+: Gracias a ella se mantiene el potencial de membrana que capacita a la célula, entre otras cosas, para captar estímulos externos. Dicho potencial se debe a que el interior celular tiene una alta concentración de K+ y una baja concentración de Na+ respecto al medio externo. La bomba de Na+ - K+ tiene actividad enzimática ATP-asa, y mantiene el gradiente porque es capaz de expulsar Na+ e introducir K+.
- 59. ENDOCITOSIS Y EXOCITOSIS Las células intercambian macromoléculas con el medio, las cuales pueden penetrar en la célula (endocitosis) o ser expulsadas de la misma (exocitosis). • Endocitosis: La endocitosis es el conjunto de procesos por medio de los cuales la célula incorpora moléculas de gran tamaño, y partículas sólidas. Consiste en que una porción de la membrana plasmática envuelve a dicho material, que queda encerrado en una vesícula endocítica, derivada de la membrana. Es un proceso activo, en el sentido de que las células consumen energía en su realización, y puede ser de tres tipos: fagocitosis, pinocitosis y endocitosis mediada por receptor. • La exocitosis: la exocitosis es el conjunto de procesos por los cuales la célula elimina al exterior moléculas de gran tamaño. Tales macromoléculas se engloban en vesículas citoplasmáticas que se desplazan hacia la superficie celular, se funden las membranas plasmática y vesicular y el contenido de la vesícula se vacía al espacio extracelular.
- 60. TRANSPORTE EN MASA Hay dos tipos de transporte en masa a través de la membrana que requieren de la acción del citoesqueleto (microfilamentos de actina) y consumo de energía: • La endocitosis: supone la entrada de nutrientes al interior de la célula. Distinguimos tres tipos: • Fagocitosis (mediante la proyección de pseudópodos • Pinocitosis (moléculas pequeñas y fluidas) • Endocitosis mediada por receptor. • La exocitosis: supone la salida de sustancias de la célula al exterior.
- 61. FAGOCITOSIS Los pseudópodos son proyecciones de membrana plasmática y citoplasma mediadas por los microfilamentos del citoesqueleto. Las células los utilizan para desplazarse sobre la superficie de un sustrato y para capturar el alimento mediante fagocitosis. Son típicos en algunos protozoos como las amebas y los foraminíferos y en algunos glóbulos blancos como los monocitos, neutrófilos y los macrófagos que capturan a sus presas o el alimento y lo fagocitan. Ameba en fagocitosis
- 62. Endocitosismediadaporreceptor LA ENDOCITOSIS MEDIADA POR RECEPTOR Implica la selección de macromoléculas específicas, especialmente importantes para el organismo, para lo cual se produce la asociación de las mismas a puntos determinados de la membrana. La cara externa de la membrana contiene receptores específicos de la molécula en cuestión, mientras que su cara interna está asociada a una proteína filamentosa (la clatrina). Las vesículas así formadas se denominan vesículas revestidas. El revestimiento se pierde una vez que se internaliza la vesícula, que posteriormente se unirá a distintos compartimentos celulares.
- 63. La exocitosis supone la salida de sustancias de la célula al exterior.
- 64. Resumen de los diferentes tipos de transporte a través de las membranas
- 65. EL CITOPLASMA: CITOSOL + ORGÁNULOS El citoplasma está constituido por el hialoplasma o citosol y los orgánulos limitados por membrana. • El hialoplasma o citosol es, pues, la parte del citoplasma que constituye el espacio comprendido entre los orgánulos membranosos. • Composición: El citosol está constituido mayoritariamente por agua; además, contiene una gran variedad de sustancias que intervienen en el metabolismo celular: proteínas, RNA de distintos tipos, aminoácidos, glúcidos, nucleótidos, lípidos e iones de distinta naturaleza. En su composición, destaca la gran abundancia de proteínas, ya que a menudo entre el 25 y el 50 % de las proteínas celulares forma parte del hialoplasma. Entre las proteínas, se encuentran miles de enzimas y otros tipos de proteínas que, como veremos un poco más adelante, forman estructuras organizadas (citoesqueleto). Esta abundancia de proteínas en dispersión coloidal, pero altamente organizada puede adquirir dos estados físicos: sol (más fluido) y gel (más viscoso). Estos estados son interconvertibles. En el citosol se encuentran también inclusiones, principalmente de glucógeno o de lípidos, no rodeadas de membrana. • Funciones del citosol: • Reserva de materiales con función energética: glucosa en disolución, partículas de glucógeno y gotitas lipídicas. • Medio donde se localizan numerosas rutas metabólicas, tanto anabólicas como catabólicas. No es un medio caótico puesto que el citoesqueleto facilita la organización de las enzimas que participan en las diferentes rutas metabólicas.
- 66. Inclusiones de Glucógeno Inclusiones de triglicéridos En el citosol se encuentran inclusiones no rodeadas de membrana y que tienen una función energética: • Inclusiones de glucógeno • Inclusiones de lípidos (triglicéridos)
- 67. El Retículo Endoplasmático es un conjunto de cavidades, túbulos y vesículas conectadas entre sí y rodeadas por una única membrana, que se prolonga formando la envoltura nuclear. El espacio que queda limitado en el interior se denomina lumen. Se distinguen dos zonas bien diferenciadas: el retículo endoplasmático rugoso (RER); y el retículo endoplasmático liso (REL).
- 68. RETÍCULO ENDOPLASMÁTICO RUGOSO Su función viene determinada por la presencia de ribosomas. Estos ribosomas proceden del citosol y su incorporación al retículo depende de la asociación entre el ribosoma y el ARNm. Si el ARNm que se acopla a los ribosomas codifica para una proteína que debe utilizarse en el citosol, el conjunto formado por el ribosoma y su ARNm permanece en el citosol (polirribosomas). Si el ARNm codifica para una proteína que debe ser procesada en el retículo endoplasmático, el conjunto ARNm y el ribosoma se dirigen a la membrana del retículo. Al mismo tiempo que se va sintetizando, la proteína va siendo transferida al lumen del retículo. Una vez allí, las proteínas son modificadas químicamente y almacenadas. Siguen este proceso las proteínas de las membranas plasmáticas y también las que salen al exterior de la célula e intervienen en la composición del glicocaliz y de la matriz extracelular, y en general las que serán vertidas al exterior de la célula por exocitosis.
- 70. RIBOSOMAS Los ribosomas se encuentran libres en el citosol o unidos al Retículo Endoplasmático Rugoso. Están constituidos por dos subunidades, la grande y la pequeña. Participan en la síntesis de proteínas.
- 71. RIBOSOMAS Los ribosomas eucariotas son 80S y están constituidos por dos subunidades: • Grande (60S). Constituida por tres ARNs (5,8S, 28S y 5S) y 45 proteínas • Pequeña (40S). Constituida por un ARN 18S y 33 proteínas. Pueden estar libres en el citosol constituyendo polirribosomas que traducen la información del ARNm en cadenas polipeptídicas. Polirribosomas procariotas en la fase de traducción
- 72. RIBOSOMAS Los ribosomas eucariotas son 80S y están constituidos por dos subunidades: • Grande (60S). Constituida por tres ARNs (5,8S, 28S y 5S) y 45 proteínas • Pequeña (40S). Constituida por un ARN 18S y 33 proteínas. Pueden estar libres en el citosol constituyendo polirribosomas que traducen la información del ARNm en cadenas polipeptídicas, como vemos en estos dibujos, o ligados al RER como veremos en la diapositiva siguiente. Polirribosomas procariotas en la fase de traducción
- 73. Si el ARNm codifica para una proteína que debe ser procesada en el retículo endoplasmático, el conjunto ARNm y el ribosoma se dirigen a la membrana del retículo. Al mismo tiempo que se va sintetizando, la proteína va siendo transferida al lumen del retículo. Una vez allí, las proteínas son modificadas químicamente y almacenadas. Siguen este proceso las proteínas de las membranas plasmáticas y también las que salen al exterior de la célula e intervienen en la composición del glicocaliz y de la matriz extracelular, y en general las que serán vertidas al exterior de la célula por exocitosis.
- 75. RETÍCULO ENDOPLASMÁTICO LISO • Se caracteriza por sacos y túbulos con membrana que carece de ribosomas adheridos. Desempeña las siguientes funciones: • Síntesis de fosfolípidos y colesterol. Estas sustancias se incorporan a las membranas de las células o intervienen en la síntesis de otros compuestos. • Destoxificación. Se encarga de procesar sustancias tóxicas procedentes del exterior de la célula. • En las células musculares actúa como almacén y regulador de la concentración de calcio en el citosol, ya que este ion es decisivo en los procesos de contracción y relajación muscular.
- 77. APARATO DE GOLGI El complejo de Golgi, o aparato de Golgi, está formado por los dictiosomas, un conjunto de cisternas o sáculos apilados, como platos, y relacionados entre sí, que aparecen rodeados de pequeñas vesículas membranosas. Este orgánulo presenta polaridad, es decir, en los dictiosomas se diferencian dos caras con distinta estructura y función: • La cara cis o de formación, por lo general más próxima al núcleo de la célula y constituida por cisternas convexas conectadas con el retículo endoplasmático rugoso a través de vesículas de transporte (de hecho, su composición química es muy semejante a la de éste). • La cara trans o cara de maduración, orientada hacia la membrana plasmática (a la que se asemeja en su composición química), y en la cual las cisternas presentan un grosor mayor; a partir de estas cisternas se originan numerosas vesículas de secreción. Entre una cara y otra se dispone un número variable de vesículas de transición. de transporte de secreción
- 79. FUNCIONES DEL APARATO DE GOLGI • Modificación de proteínas sintetizadas en el RER. • Secreción de proteínas. • Participa en la formación de la pared celular vegetal. • Participa en la formación del glicocaliz en las células animales. • Participa en el tránsito de lípidos por la célula.
- 80. El APARATO DE GOLGI • Las proteínas y los lípidos producidos en el retículo endoplasmático pasan a través de vesículas de transporte a la cara cis del aparato de Golgi. Van madurando a lo largo de los dictiosomas de transición hasta la cara trans. El proceso de maduración consiste en añadir o modificar la parte glucídica o activar a la enzima que está inactiva. Por último, en la cara trans se forman las vesículas de secreción, que se fusionan con la membrana plasmática y liberan su contenido al exterior. • Algunas vesículas de secreción dan lugar a los lisosomas. Las sustancias que se dirigen a los lisosomas son enzimas sintetizadas en el RER que han sido almacenadas en el aparato de Golgi sin haber experimentado ninguna transformación. Estos enzimas son imprescindibles para la actividad hidrolítica de estos orgánulos (digestión celular).
- 81. En el aparato de Golgi se producen las proteínas que: • Van a formar parte de la membrana plasmática • Van a formar parte de la membrana del Retículo Endoplasmático • Van a ser secretadas al exterior de la célula • Van a formar los lisosomas. Existen diferentes señales moleculares en las vesículas de transporte que serán reconocidas específicamente en sus lugares de destino.
- 82. El proceso de síntesis y secreción de proteínas a través de los distintos compartimentos celulares se ha observado en ciertas células, como por ejemplo, las células secretoras del páncreas, especializadas en la secreción de enzimas digestivas. El método utilizado constituye una combinación de técnicas autorradiográficas y MET, para el que se emplean aminoácidos con tritio (H3).
- 83. A los cinco minutos el marcaje radiactivo aparece en el RER; a los 15 minutos el marcaje radiactivo aparece en el aparato de Golgi; y a partir de los 30 minutos el marcaje radiactivo va apareciendo en las vesículas de secreción.
- 84. TRÁFICO EN EL SISTEMA DE ENDOMEMBRANAS. Las vesículas que transportan lípidos y proteínas pueden seguir varias rutas desde el RE y el complejo de Golgi, dando lugar a vesículas de secreción, endosomas y lisosomas. 1. Las proteínas se sintetizan en los ribosomas unidos a la cara citosólica del RE rugoso. 2. Las etapas iniciales de la glicosilación tienen lugar en la luz del RE. Las vesículas de transición llevan hacia la RCG a los lípidos y glicoproteínas recién sintetizados. 3. Los lípidos y proteínas se desplazan por el dictiosoma gracias a vesículas o a cisternas que van madurando. Desde la RTG parten las que serán las vesículas de secreción 4. y las que formarán parte de los endosomas, 5. en función de su contenido proteico. Las vesículas de secreción se dirigen hacia la membrana plasmática, donde liberan, su contenido por exocitosis, bien constitutiva, bien como respuesta a una señal. 6. La célula toma por endocitosis proteínas y otros materiales, fabricando así vesículas de endocitosis, que se fusionan con los endosomas tempranos. 7. Los componentes no destinados a la digestión que sigue a la endocitosis, son reciclados hacia la membrana plasmática. 8. Los endosomas tempranos, con el material destinado a la digestión, maduran dando lugar, primero a los endosomas tardíos, y luego a los lisosomas. 9. El tráfico retrógrado permite el retorno de proteínas específicas a sus compartimentos correspondientes.
- 85. LOS LISOSOMAS • Los lisosomas son pequeñas vesículas, limitadas por membrana, que contienen una gran variedad de enzimas hidrolíticas implicadas en los procesos de digestión celular y que se forman en el Aparato de Golgi. • Las enzimas lisosómicas son hidrolasas, cuya actividad óptima tiene lugar a pH ácido (alrededor de pH 5)
- 86. LOS LISOSOMAS Los lisosomas son pequeñas vesículas, limitadas por membrana, que contienen una gran variedad de enzimas hidrolíticas implicadas en los procesos de digestión celular y que se forman en el Aparato de Golgi. • Las enzimas lisosómicas son hidrolasas, cuya actividad óptima tiene lugar a pH ácido (alrededor de pH 5). • La membrana lisosómica posee una importancia extraordinaria para el correcto funcionamiento del orgánulo, ya que contiene proteínas de transporte específicas que mantienen el pH ácido y otras proteínas implicadas en el transporte de determinadas sustancias hacia el interior del lisosoma o de los productos de la digestión hacia el citoplasma celular. • El pH interno se mantiene en un valor medio de 5 gracias a una enzima ATPasa de membrana que bombea H+ hacia el interior a expensas de la hidrólisis de ATP. • Las proteínas de la cara interna de la membrana lisosómica están altamente glucosiladas, lo cual les sirve de protección frente a sus propias enzimas y la acidez del medio.
- 87. LOS LISOSOMAS Tipos de lisosomas: • Lisosomas primarios: de reciente formación, proceden del aparato de Golgi y contienen diversas enzimas hidrolíticas. Se caracterizan por poseer un contenido homogéneo. • Lisosomas secundarios: en ellos tienen lugar procesos activos de digestión celular. Presentan un contenido heterogéneo que depende del tipo de nutrición de la célula. Funciones de los lisosomas. Los lisosomas participan activamente en los procesos de digestión celular. Dependiendo de la función que desempeñen en dichos procesos, se clasifican en: • Fagolisosomas (heterofagia) • Autofagolisosomas (autofagia) Lisosomas primarios (flechas rojas) y lisosomas secundarios
- 88. LOS LISOSOMAS Los lisosomas participan activamente en los procesos de digestión celular. Dependiendo de la función que desempeñen en dichos procesos, se clasifican en fagolisosomas (heterofagia) y autofagolisosomas (autofagia) • Fagolisosomas: (vacuolas digestivas o heterolisosomas). Se trata de orgánulos formados a partir de la unión de un lisosoma primario con una vacuola fagocítica, que contiene partículas alimenticias de naturaleza muy variada según el tipo de célula. Son abundantes en las amebas y en los macrófagos (leucocitos especializados en la fagocitosis de microorganismos patógenos). • Autofagolisosomas: En este caso, los lisosomas se fusionan con vacuolas autofagocíticas para eliminar restos celulares, como orgánulos dañados por sustancias tóxicas o que deben ser reciclados. Una vez finalizada la digestión celular, en los lisosomas secundarios quedan restos que no pueden ser aprovechados por la célula y son excretados al exterior, aunque en ciertos casos permanecen en la célula como cuerpos residuales.
- 94. LOS PEROXISOMAS Son parecidos a los lisosomas, diferenciándose de estos en que contienen enzimas que degradan los ácidos grasos y los aminoácidos. Como estos procesos generan peróxidos, contienen también catalasa, enzima que descompone los peróxidos y en particular el H2O2 en H2O y O2.
- 96. LOS GLIOXISOMAS Los glioxisomas son una clase de peroxisomas que sólo se encuentran en las células de los vegetales. Su nombre deriva de que poseen las enzimas responsables del ciclo del ácido glioxílico, una variante del ciclo de Krebs, que permite sintetizar glúcidos a partir de lípidos. Esto resulta esencial para las semillas en germinación, ya que les permite, .a partir de sus reservas lipídicas, sintetizar glucosa, única molécula que admite el embrión, hasta que el nuevo vegetal pueda extender sus hojas y realizar la fotosíntesis
- 98. LAS VACUOLAS Son cavidades intracitoplasmáticas, limitadas por una fina membrana o tonoplasto, donde se acumula agua con diversas sustancias disueltas. Aunque también existen en las células animales, donde son numerosas y pequeñas, están mucho más desarrolladas en las células vegetales donde su conjunto recibe el nombre de vacuoma. Su tamaño y forma varían según el estado de desarrollo y el estado metabólico de la célula: en las células meristemáticas, es decir, sin diferenciar, suelen ser numerosas y pequeñas pero, a medida que la célula crece y se hace más vieja, las vacuolas aumentan de tamaño y se unen gradualmente. Así, muchas células adultas presentan sólo una gran vacuola que ocupa la parte central de la célula, mientras que el citoplasma queda restringido a una estrecha banda junto a la membrana.
- 99. FUNCIONES DE LAS VACUOLAS Las vacuolas proceden de vesículas derivadas del RE y del Aparato de Golgi, y realizan las siguientes funciones: • Acumular agua: con ello se consigue un aumento del tamaño celular, que resulta muy económico. • Almacenar sustancias muy diversas: de reserva, productos de desecho tóxicos, sales, pigmentos y esencias para atraer a los insectos polinizadores, sustancias nocivas para evitar que sean depredadas. • Pueden actuar con funciones análogas a los lisosomas cuando contienen enzimas hidrolíticas. • Contribuyen a la homeostasis del interior celular. • En otras células no vegetales existen vacuolas contráctiles. Son características de microorganismos que viven en medios hipotónicos respecto del interior celular, como diversos grupos de protozoos.
- 101. La Teoría endosimbiótica postula que las mitocondrias y los cloroplastos evolucionaron a partir de bacterias capaces de realizar la respiración celular y la fotosíntesis respectivamente.
- 102. MITOCONDRIAS Las mitocondrias son orgánulos comunes tanto en células animales como vegetales. En ellas se realiza el metabolismo respiratorio aeróbico, cuya finalidad es la obtención de energía. Las mitocondrias presentan una forma y tamaños variables, aunque por lo general son cilíndricas o alargadas y con los extremos redondeados; su diámetro oscila entre 0,5 y 1 µm. Aparecen en número variable según el tipo de célula. Son especialmente abundantes en aquellas células que requieren un elevado aporte energético. Estructura y composición de las mitocondrias: • Membrana mitocondrial externa • Espacio intermembrana • Membrana mitocondrial interna que forma crestas mitocondriales • Matriz mitocondrial (con ribosomas y el Cromosoma mitocondrial)
- 103. ESTRUCTURA DE LAS MITOCONDRIAS • Membrana mitocondrial externa. Contiene un reducido número de proteínas con actividad enzimática y porinas (proteínas intermembranales que permiten el paso de moléculas pequeñas) en abundancia. • Espacio intermembrana. Se localiza entre ambas membranas mitocondriales y está ocupado por una matriz de composición semejante a la del citosol. • Membrana mitocondrial interna. Posee la estructura trilaminar típica del resto de las membranas celulares y presenta invaginaciones o crestas mitocondriales que se introducen en la matriz. Las crestas pueden ser aplanadas (vesiculares) o tubulares y, por lo general, se disponen perpendicularmente al eje mayor de la mitocondria. En cuanto a su composición, esta membrana carece de colesterol y es más impermeable a los iones que la membrana externa. En ella se encuentran las cadenas de transporte electrónico y enzimas como la ATPasa. • Matriz mitocondrial. Contiene ADN mitocondrial circular, ARN y ribosomas 70S como las bacterias. Incluye, además, diversas enzimas responsables del ciclo de Krebs, y transportadores de electrones como el NADH.
- 105. MITOCONDRIAS La función de las mitocondrias consiste en obtener energía para la célula. Esta función se halla compartimentalizada en la matriz mitocondrial y la membrana mitocondrial interna, en las que tienen lugar distintos procesos. En la matriz mitocondrial se produce: • La β-oxidación de los ácidos grasos. • La descarboxilación del ácido pirúvico (originado a partir de los glúcidos por glucólisis), procedentes del citosol. Se generan acetil-CoA y moléculas reducidas (NADH+H+ y FADH2). • El ciclo de Krebs. El acetil-CoA es oxidado completamente a CO2 y se obtienen intermediarios metabólicos y moléculas reducidas (NADH+H+ y FADH2). • La síntesis de proteínas mitocondriales a expensas de la maquinaria replicativa y del ADN mitocondriales.
- 106. FOSFORILACIÓN OXIDATIVA EN LAS MITOCONDRIAS En la membrana mitocondrial interna que forma crestas mitocondriales se realiza la fosforilación oxidativa. El NADH+H+ y el FADH2 (moléculas con poder reductor) originados en la matriz son los donadores de electrones a la cadena respiratoria (complejo I y II respectivamente) en la que el último aceptor es el O2, que con protones da lugar a H2O. Los complejos I, III y IV de la cadena respiratoria realizan un bombeo de protones (transporte activo) al espacio intermembrana que genera un gradiente electroquímico de H+. Este gradiente es aprovechado por la ATPasa de la membrana interna que actúa como canal de protones y promueve la fosforilación del ADP para sintetizar ATP (fosforilación oxidativa). ATPasa I II III IV
- 107. FOSFORILACIÓN OXIDATIVA EN LAS MITOCONDRIAS En la membrana mitocondrial interna que forma crestas mitocondriales se realiza la fosforilación oxidativa. El NADH+H+ y el FADH2 (moléculas con poder reductor) originados en la matriz son los donadores de electrones a la cadena respiratoria (complejo I y II respectivamente) en la que el último aceptor es el O2, que con protones da lugar a H2O. Los complejos I, III y IV de la cadena respiratoria realizan un bombeo de protones (transporte activo) al espacio intermembrana que genera un gradiente electroquímico de H+. Este gradiente es aprovechado por la ATPasa de la membrana interna que actúa como canal de protones y promueve la fosforilación del ADP para sintetizar ATP (fosforilación oxidativa).
- 109. DIVISIÓN DE LAS MITOCONDRIAS Las mitocondrias se dividen en el interior de las células eucariotas por bipartición o fisión binaria como las bacterias.
- 110. o plastos
- 111. CLOROPLASTOS En los cloroplastos de las células vegetales se realiza la fotosíntesis oxigénica en condiciones de iluminación. La fotosíntesis oxigénica es un proceso metabólico por el cual la célula obtiene energía a partir de la luz y la convierte en energía química, que es utilizada posteriormente para la fijación de CO2 en moléculas orgánicas. Los cloroplastos se encuentran en número y forma variable (oval, estrellada, acintada, en copa, helicoidal, etc), principalmente en los protoctistas. Las formas microscópicas suelen contener uno o dos cloroplastos de gran tamaño, o bien cloroplastos numerosos más pequeños, distribuidos por lo general en la periferia. En los vegetales superiores, su número oscila entre 20 y 40 por célula, dependiendo del tejido en el que se encuentren (por ejemplo, son muy abundantes en el parénquima clorofílico), y la forma más frecuente es la ovoide. Los cloroplastos son más grandes que las mitocondrias: su tamaño oscila entre 2 y 10 µm. La teoría endosimbiótica nos explica su origen evolutivo a partir de bacterias fotosintéticas. Se dividen por bipartición o fisión binaria.
- 112. Dibujo esquemático y fotografía al MET de un cloroplasto ESTRUCTURA DE LOS CLOROPLASTOS • Envoltura: Tanto la membrana externa como la interna poseen una estructura continua y delimitan completamente el cloroplasto. Ambas están separadas por el espacio intermembrana. • Estroma: Es la matriz del cloroplasto, delimitada por la membrana plastidial interna que rodea a los grana. El estroma contiene ADN circular de doble hélice, ribosomas 70S como los de las bacterias, gránulos de almidón e inclusiones lipídicas. En él se localizan la rubisco y otras enzimas implicadas en la fijación del CO2 en el ciclo de Calvin. • Tilacoides y grana: En el interior del estroma se pueden visualizar los tilacoides, una serie de sáculos membranosos que se disponen paralelamente al eje mayor del cloroplasto. Algunos tilacoides se apilan formando grupos, los grana (singular granum), mientras que otros, de mayor longitud, atraviesan el estroma y conectan los distintos grana entre sí. Tilacoide Estroma
- 113. LOS TILACOIDES Las membranas tilacoidales presentan una composición diferente a las que integran la envoltura y contienen alrededor de un 12% de pigmentos, principalmente clorofilas, carotenos y otros pigmentos accesorios. En estas membranas se localizan los fotosistemas (centros de reacción fotosintéticos) y los pigmentos antena, así como las cadenas de transporte electrónico y las ATPasas implicadas en el proceso de fotofosforilación.
- 114. LA FOTOSÍNTESIS Los cloroplastos son los orgánulos encargados de realizar la fotosíntesis oxigénica. En este proceso metabólico, en el que el agua actúa como donador de electrones y se genera oxígeno, la célula utiliza la luz como fuente de energía, y el CO2, como fuente de carbono. En este proceso se distinguen dos fases: • Fase luminosa: En la membrana tilacoidal se producen las reacciones de conversión de la energía lumínica en energía química (ATP) y se genera poder reductor (NADPH+H+), para lo cual son imprescindibles la luz, la presencia de pigmentos fotosintéticos y el agua. Se libera O2. • Fase oscura: En el estroma tiene lugar la fijación del CO2 en moléculas orgánicas (ciclo de Calvin) y su almacenamiento en forma de polisacáridos de reserva, por lo general almidón. Se requiere el ATP y el poder reductor en forma de NADPH producido en la fase luminosa. Las reacciones del ciclo de Calvin son independientes de la luz, de ahí la denominación de fase oscura. La enzima más importante del ciclo es la Rubisco.
- 115. LA FOTOSÍNTESIS • Fase luminosa: En la membrana tilacoidal se producen las reacciones de conversión de la energía lumínica en energía química (ATP) y se genera poder reductor (NADPH+H+), para lo cual son imprescindibles la luz, la presencia de pigmentos fotosintéticos y el agua. Se libera oxígeno.
- 116. LA FOTOSÍNTESIS • Fase oscura: En el estroma tiene lugar la fijación del CO2 en moléculas orgánicas (ciclo de Calvin) y su almacenamiento en forma de polisacáridos de reserva, por lo general almidón. Se requiere el ATP y el poder reductor en forma de NADPH producido en la fase luminosa. Las reacciones del ciclo de Calvin son independientes de la luz, de ahí la denominación de fase oscura. La enzima más importante del ciclo es la Rubisco.
- 117. Núcleo interfásico EL NÚCLEO • En 1830, Brown estableció la constancia del núcleo en todas las células eucariotas. • Este orgánulo contiene la información genética de la célula, por lo que es responsable de la transmisión del mensaje genético a través de las generaciones y de dar a la célula las instrucciones para su crecimiento y desarrollo. • El núcleo se presenta en dos estados diferentes que dependen de la fase del ciclo celular: interfase y división celular. • El núcleo interfásico coincide con las fases del ciclo celular en que no se divide. Este núcleo interfásico está constituido por una doble membrana, o envoltura nuclear, y un interior formado por el nucleoplasma y el material genético: la cromatina y el nucleolo. • El núcleo interfásico se caracteriza por el hecho de que su forma, tamaño, posición y número son diferentes, según el tipo celular y el estado de desarrollo de la célula. Su forma, por lo general, es esférica, aunque varía notablemente de un tipo celular a otro. El tamaño suele ser de 2 a 25 µm. de diámetro, aumentando considerablemente antes de realizarse la división celular, debido a la duplicación del material genético. La posición depende del tipo de actividad celular. Suele haber un núcleo por célula, pero hay numerosas excepciones.
- 118. Núcleo interfásico ENVOLTURA NUCLEAR • El núcleo se encuentra rodeado por una membrana doble: la membrana nuclear externa y la membrana nuclear interna, separadas por un espacio perinuclear que oscila entre 25 y 40 nm. La membrana nuclear externa está conectada y se continúa con el RER. • Ambas membranas se comunican por medio de poros nucleares (80 nm.), a través de los cuales se produce el intercambio y transporte de moléculas grandes, como el ARN las proteínas entre el nucleoplasma y el citoplasma.
- 119. Núcleo interfásico • El nucleoplasma del núcleo se comunica con el citosol por medio de poros nucleares (80 nm.), a través de los cuales se produce el intercambio y transporte de moléculas grandes, como el ARN y las proteínas. • Los poros, más abundantes en las células no diferenciadas o muy activas, constituyen un complejo proteico característico. Cada poro está rodeado por un anillo integrado por ocho bloques proteicos dispuestos en forma octogonal. Cada bloque, a su vez, consta de más de cien proteínas distintas. Las proteínas del anillo mantienen la morfología de los poros, sirven de anclaje a la membrana nuclear y conectan el poro con una masa densa situada en el centro del mismo a través de finas fibrillas. • Entre el nucleoplasma y la membrana nuclear interna se observa, por último, una lámina nuclear fibrosa constituida por filamentos intermedios. Su función parece ser la regulación de las interacciones entre la cromatina y la envoltura nuclear, la organización de ésta, así como su desaparición y nueva formación durante la mitosis. Poros nucleares Complejo del Poro Nuclear
- 120. Lámina nuclear
- 122. El ADN de los eucariotas se une a proteínas de dos tipos: histonas y proteínas cromosómicas no histónicas. Estas últimas incluyen miles de proteínas en su mayor parte enzimas que intervienen en la duplicación y transcripción del ADN. Cromatina: La cromatina está compuesta por ADN plegado, asociado a proteínas básicas estructurales y reguladoras: Las Histonas. En la cromatina del núcleo interfásico, la doble hélice de ADN se asocia a histonas y forma complejos denominados nucleosomas. Se podría decir que el nucleosoma es la unidad básica estructural de la cromatina. alrededor del nucleosoma se enrolla una cadena de 140 pares de bases de ADN, y entre cada unidad hay un fragmento de ADN. Las histonas son proteínas básicas que contienen una elevada proporción de aminoácidos básicos Hay dos grupos: • Histonas nucleosómicas: componen un complejo proteico discoidal alrededor del cual se enrolla el ADN (dos vueltas de la doble hélice de ADN por nucleosoma). Cada uno de estos discos es un octámero formado por dos copias de cada histona (H2A, H2B, H3 y H4). Esta estructura constituye el nucleosoma. • Histona H1: une los complejos nucleosómicos y es la responsable del plegamiento helicoidal de la fibra elemental de cromatina. La cadena de nucleosomas es la fibra nucleosómica o unidad elemental de la cromatina, de unos 10 nm de grosor.
- 123. El ADN de los eucariotas se une a proteínas de dos tipos: histonas y proteínas cromosómicas no histónicas. Estas últimas incluyen miles de proteínas en su mayor parte enzimas que intervienen en la duplicación y transcripción del ADN. Cromatina: La cromatina está compuesta por ADN plegado, asociado a proteínas básicas estructurales y reguladoras: Las Histonas. En la cromatina del núcleo interfásico, la doble hélice de ADN se asocia a histonas y forma complejos denominados nucleosomas. Se podría decir que el nucleosoma es la unidad básica estructural de la cromatina. alrededor del nucleosoma se enrolla una cadena de 146 pares de bases de ADN, y entre cada unidad hay un fragmento de ADN libre de unos 50 pares de bases. Las histonas son proteínas básicas que contienen una elevada proporción de aminoácidos básicos Hay dos grupos: • Histonas nucleosómicas: componen un complejo proteico discoidal alrededor del cual se enrolla el ADN (dos vueltas de la doble hélice de ADN por nucleosoma). Cada uno de estos discos es un octámero formado por dos copias de cada histona (H2A, H2B, H3 y H4). Esta estructura constituye el nucleosoma. • Histona H1: une los complejos nucleosómicos y es la responsable del plegamiento helicoidal de la fibra elemental de cromatina. La cadena de nucleosomas es la fibra nucleosómica o unidad elemental de la cromatina, de unos 10 nm de grosor.
- 124. Las fibras complejas de cromatina se originarían por el superenrollamiento de la cadena de nucleosomas según una disposición regular, en la que la histona H1 desempeña un papel esencial. Según la hipótesis más aceptada hoy día, la cadena de nucleosomas se enrollaría helicoidalmente formando un solenoide o fibra de 30 nm., que contendría seis nucleosomas por cada vuelta de hélice. Esta estructura estaría estabilizada por las histonas H1, dispuestas en el núcleo del nucleosoma, que interaccionan con los fragmentos de ADN libre o internucleosómico. A su vez, las fibras complejas de 30 nm. se hallan plegadas en el núcleo interfásico en forma de bucles radiales, que alcanzarán otros niveles de compactación y enrollamientos sucesivos en el núcleo en división hasta llegar a constituir los cromosomas metafásicos.
- 125. Fibra nucleosómica de 10 nm. Vista al MET. Solenoide o fibra de 30 nm. Vista al MET.
- 126. Las fibras complejas de cromatina se originarían por el superenrollamiento de la cadena de nucleosomas según una disposición regular, en la que la histona H1 desempeña un papel esencial. Según la hipótesis más aceptada hoy día, la cadena de nucleosomas se enrollaría helicoidalmente formando un solenoide o fibra de 30 nm., que contendría seis nucleosomas por cada vuelta de hélice. Esta estructura estaría estabilizada por las histonas H1, dispuestas en el núcleo del nucleosoma, que interaccionan con los fragmentos de ADN libre o internucleosómico. A su vez, las fibras complejas de 30 nm. se hallan plegadas en el núcleo interfásico en forma de bucles radiales, que alcanzarán otros niveles de compactación y enrollamientos sucesivos en el núcleo en división hasta llegar a constituir los cromosomas metafásicos.
- 127. De la doble hélice de 2 nm hasta el cromosoma metáfasico, formado por dos cromátidas hermanas de 700 nm, podemos observar cuatro niveles de empaquetamiento o compactación: 1. Fibra nucleosómica de 10 nm. 2. Fibra de 30 nm o Solenoide. 3. Fibra de 300 nm formada por rosetones de 6 bucles. 4. Fibra de 700 nm, Rodillo o cromátida formada por 30 rosetones.
- 128. De la doble hélice de 2 nm hasta el cromosoma metáfasico, formado por dos cromátidas hermanas de 700 nm, podemos observar cuatro niveles de empaquetamiento o compactación: 1. Fibra nucleosómica de 10 nm. 2. Fibra de 30 nm o Solenoide. 3. Fibra de 300 nm formada por rosetones de 6 bucles. 4. Fibra de 700 nm, Rodillo o cromátida formada por 30 rosetones.
- 129. Armazón central de un cromosoma y detalle de los bucles extendidos
- 130. TIPOS DE CROMATINA EN EL NÚCLEO INTERFÁSICO En el núcleo en interfase se diferencian dos tipos de cromatina: • Eucromatina: De aspecto laxo o difuso, corresponde a zonas de cromatina activas (aproximadamente, el 10 % de la cromatina celular), donde se produce la transcripción. • Heterocromatina: Se visualiza al microscopio como áreas más densas y homogéneas de cromatina altamente condensada (el 90 % del total), que corresponden a zonas inactivas que no se transcriben. Se pueden distinguir también dos tipos de heterocromatina: • Constitutiva, que aparece condensada siempre durante todo el ciclo celular y cuyo ADN no se transcribe nunca. • Facultativa, cuya condensación depende del estado de desarrollo del organismo y del tipo celular, y comprende el conjunto de genes que, de manera específica, se inactivan a lo largo de la diferenciación celular. El Nucleoplasma, también llamado carioplasma, es el medio interno acuoso donde se encuentran inmersos los demás componentes nucleares. Está integrado por proteínas, principalmente enzimas relacionadas con el metabolismo de ADN y ARN. En numerosas ocasiones se han detectado en el núcleo interfásico algunos elementos citoesqueléticos (lámina nuclear) que podrían formar una trama que interacciona con el ADN de la cromatina y con la envoltura nuclear. Euc Heter Euc Heter Nucleolo
- 131. EL NUCLEOLO • El nucleolo es una estructura esférica visible en el interior del núcleo interfásico, cuya función principal es la síntesis y ensamblaje de las subunidades ribosómicas. • Algunas células presentan más de un nucleolo. • El nucleolo está constituido por proteínas y ácidos nucleicos, ADN (1-3 %) y ARN, este último en una proporción mayor (10-30 %). • Esta estructura interviene activamente en la transcripción de ARNr y su desarrollo en la célula parece guardar relación con la formación de ribosomas para la síntesis de proteínas. El ADN nucleolar que codifica para la síntesis de ARNr y puede formar parte de cromosomas distintos constituye el organizador nucleolar. Los ARNr recién sintetizados maduran y se ensamblan con las proteínas ribosómicas importadas del citoplasma, originando las subunidades de 60S y 40S que emigran al citoplasma, donde se ensamblan a su vez en presencia de ARNm. Euc Heter Euc Heter Nucleolo
- 132. EL NUCLEOLO Los nucleolos están constituidos por una parte amorfa o nucleoplasma y una parte densa, compuesta, a su vez, por una zona granular, con ribonucleoproteínas, y una zona fibrilar que contiene ADN y ARN. La zona fibrilar podría corresponder a áreas de transcripción activa (organizadores nucleolares), mientras que en la sección granular se produce la maduración de las subunidades ribosómicas, que son exportadas por separado al citoplasma, donde se unirán para formar los ribosomas completos.
- 133. En el núcleo mitótico observamos cromosomas al microscopio óptico. El tamaño de los cromosomas y el número es variable y depende de la especie. Los cromosomas son estructuras cilíndricas que representan el grado más elevado de empaquetamiento del ADN y, por tanto, de la cromatina en la célula. Durante la metafase, cada cromosoma aparece constituido por dos brazos o cromátidas iguales, unidos por el centrómero. Los centrómeros reciben también el nombre de constricciones primarias e incluyen los cinetocoros, placas de naturaleza proteica a las que están conectados los microtúbulos cromosómicos del huso mitótico. Según la posición del centrómero, los cromosomas se clasifican en: • Cromosomas metacéntricos: los brazos del cromosoma son iguales, el centrómero está en el centro del cromosoma. • Cromosomas submetacéntricos: el centrómero aparece algo desplazada y da lugar a dos brazos desiguales. • Cromosomas acrocéntricos: El centrómero está muy desplazado y los brazos que se forman son muy desiguales. • Cromosomas telocéntricos: presentan el centrómero en el extremo del cromosoma, de modo que contienen un brazo de gran longitud y otro prácticamente indistinguible. Los extremos de las cromátidas se denominan telómeros y en ellos se encuentran secuencias repetitivas de ADN que tienen como función evitar la pérdida de información genética en la replicación y la fusión de cromosomas alterados (mantiene la individualidad de los cromosomas). Los satélites se unen a constricciones secundarias de tamaño variable. Algunas de estas constricciones secundarias contienen el organizador nucleolar que originará los nucleolos en la telofase al final de la mitosis. Se trata de una zona del cromosoma en la que están los genes que codifican para los ARN ribosómicos, por lo que es indispensable para que se formen nuevos ribosomas.
- 134. CARIOTIPO Y CARIOGRAMA Cariotipo es el conjunto de rasgos característicos de los cromosomas de cada especie (tamaño, forma, etc.) Cada cromosoma o pareja de cromosomas se designa con un número, excepto los cromosomas sexuales, que se denominan con las letras X e Y. La representación gráfica de los cromosomas homólogos, ordenados de mayor a menor tamaño, se denomina Cariograma.
- 135. CITOESQUELETO El citoesqueleto constituye una red de filamentos proteicos, compleja e interconectada, responsable del mantenimiento y los cambios de la forma celular, el movimiento y posicionamiento de los orgánulos, y la división y motilidad de la célula. El citoesqueleto característico de la arquitectura celular eucariota está formado por tres tipos de filamentos: • Los microtúbulos. • Los filamentos intermedios. • Los microfilamentos de actina.
- 138. MICROTÚBULOS Se trata de estructuras fibrilares con un diámetro constante de 25 nm, integradas, mayoritariamente, por tubulina, una proteína globular. La tubulina se polimeriza de forma regular ensamblándose alrededor de un núcleo central hueco. Esta estructura se estabiliza por medio de una serie de proteínas asociadas que reciben el nombre de proteínas asociadas a los microtúbulos. Los dímeros de tubulina se ensamblan siguiendo una orientación, es decir, los microtúbulos presentan polaridad: tienden a crecer o polimerizarse en uno de sus extremos (+) y a desorganizarse o despolimerizarse en el extremo opuesto (-). Estos elementos citoesqueléticos se organizan a partir de regiones especializadas denominadas centros organizadores de microtúbulos o centrosomas, que con frecuencia contienen en el centro un par de centriolos constituidos por nueve tripletes de microtúbulos.
- 139. MICROTÚBULOS Se trata de estructuras fibrilares con un diámetro constante de 25 nm, integradas, mayoritariamente, por tubulina, una proteína globular. La tubulina se polimeriza de forma regular ensamblándose alrededor de un núcleo central hueco. Esta estructura se estabiliza por medio de una serie de proteínas asociadas que reciben el nombre de proteínas asociadas a los microtúbulos. Los dímeros de tubulina se ensamblan siguiendo una orientación, es decir, los microtúbulos presentan polaridad: tienden a crecer o polimerizarse en uno de sus extremos (+) y a desorganizarse o despolimerizarse en el extremo opuesto (-). Estos elementos citoesqueléticos se organizan a partir de regiones especializadas denominadas centros organizadores de microtúbulos o centrosomas, que con frecuencia contienen en el centro un par de centriolos constituidos por nueve tripletes de microtúbulos.
- 140. CENTRIOLOS Y FUNCIONES DE LOS MICROTÚBULOS En las células eucariotas animales, cerca del núcleo, aparecen dos centriolos, muy próximos uno a otro y cruzados en ángulo recto. Cada centriolo está formado por nueve tripletes de microtúbulos en disposición radial constituyendo una estructura en empalizada cilíndrica y hueca. Centrosoma: Los centriolos están rodeados por un material denso o material pericentriolar, de donde irradian unos microtúbulos en todas las direcciones, que constituyen el áster. El conjunto formado por los dos centriolos, el material pericentriolar y el áster recibe el nombre de centrosoma o citocentro. Los microtúbulos se encuentran en abundancia en la mayoría de las células eucariotas y desempeñan en ellas funciones vitales: • Contribuyen al mantenimiento de la forma celular. • Participan en el transporte de orgánulos y partículas en el interior de la célula. • Constituyen los elementos estructurales fundamentales del huso mitótico durante la división nuclear. • Constituyen los elementos estructurales fundamentales de los centriolos. • Constituyen el esqueleto interno de los cilios y los flagelos, así como de sus corpúsculos basales. Áster Material Pericentriolar
- 141. Células embrionarias en división. Se aprecian los centrosomas o centros organizadores de microtúbulos que forman el huso mitótico
- 144. CILIOS Y FLAGELOS Son apéndices externos, responsables de la motilidad de las células eucariotas, que presentan una estructura común. La diferencia entre cilios y flagelos reside, fundamentalmente, en su mecanismo de movimiento. Además, los flagelos suelen ser más largos y menos numerosos que los cilios.Células ciliadas del epitelio respiratorio Espermatozoides flagelados fecundando un óvulo
- 145. ESTRUCTURA DE CILIOS Y FLAGELOS Los Cilios y Flagelos presentan la misma estructura interna en la que se distinguen los siguientes elementos: • Axonema: parte interna del tallo de un cilio o un flagelo que está formada por nueve pares de microtúbulos alrededor de una par de microtúbulos centrales (estructura 9+2). Asociada a los microtúbulos se encuentra una serie de proteínas, las más importantes de las cuales son la dineína y la nexina. Esta estructura se halla rodeada por una extensión de la membrana plasmática de la célula. Uno de los microtúbulos de cada par está incompleto. • Corpúsculo basal: se encuentra en la base del axonema y está compuesto por nueve tripletes de microtúbulos periféricos unidos radialmente a un eje central (estructura 9+0). Esta estructura es idéntica a la de los centriolos. Del extremo inferior del corpúsculo basal nacen unas fibras denominadas raíces ciliares. • Zona de transición: Situada entre el axonema y el corpúsculo basal, se observa en ella una placa basal formada por un material denso a los electrones.
- 146. FUNCIÓN DE CILIOS Y FLAGELOS • En los organismos eucariotas unicelulares y en los gametos flagelados de los organismos superiores, los cilios y flagelos están implicados en el desplazamiento de la célula. • En otros casos, como en las células epiteliales ciliadas o en los cilios de la abertura oral de algunos protistas, el movimiento de estas estructuras provoca corrientes que arrastran las partículas circundantes. Los corpúsculos basales de los cilios y los flagelos pueden encontrarse asociados a los centros organizadores de microtúbulos.
- 148. MICROFILAMENTOS Tienen un diámetro medio de 8 nm, Están formados por proteínas como la actina y la miosina, que pueden tener estructura globular o fibrosa (polimerización). Constituye parte del citoesqueleto formado por fibras de actina adosadas a la cara interna de la membrana plasmática. La consistencia de esta estructura puede cambiar según las necesidades de la célula. Desempeña las siguientes funciones: • Da forma a la membrana plasmática. • Produce deformaciones de la membrana que originan los pseudópodos. • Estabiliza la estructura de las microvellosidades en las membranas plasmáticas. • Participa en la formación de vesículas de endocitosis. • Interviene en la formación de las corrientes que se observan en el interior del citoplasma. • Forma parte del anillo contráctil que tiene lugar en la citocinesis animal. • Permite la contracción muscular en el interior de las células musculares.
- 150. FILAMENTOS INTERMEDIOS Los filamentos intermedios son componentes citoesqueléticos especialmente abundantes en las células animales. De naturaleza proteica, presentan un diámetro intermedio (8-10 nm.) entre el de los microtúbulos y el de los microfilamentos de actina, de ahí su nombre. A diferencia de los microfilamentos y microtúbulos, los filamentos intermedios son muy estables, es decir, no presentan en la célula un equilibrio de polimerización-despolimerización, y no participan en el movimiento intracitoplasmático de orgánulos y vesículas. Su función es siempre estructural, aunque su composición varía dependiendo del tipo de células: los neurofilamentos se disponen irregularmente en el citoplasma y los axones de las células neuronales; las células epiteliales contienen filamentos de queratina, que forman una red citoplasmática especialmente alrededor del núcleo; la desmina en las células musculares; También forman la lámina nuclear, situada en el núcleo y en contacto con la cara interna de la envoltura nuclear, responsable de su organización y desorganización en la mitosis.