

Recommended
More Related Content
More from Pasi Vilpas
More from Pasi Vilpas (20)
Makuaistin neurologiaa
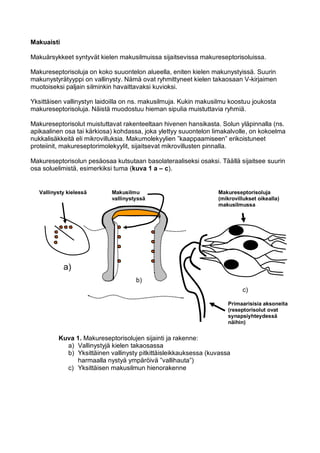
- 1. Makuaisti Makuärsykkeet syntyvät kielen makusilmuissa sijaitsevissa makureseptorisoluissa. Makureseptorisoluja on koko suuontelon alueella, eniten kielen makunystyissä. Suurin makunystyrätyyppi on vallinysty. Nämä ovat ryhmittyneet kielen takaosaan V-kirjaimen muotoiseksi paljain silminkin havaittavaksi kuvioksi. Yksittäisen vallinystyn laidoilla on ns. makusilmuja. Kukin makusilmu koostuu joukosta makureseptorisoluja. Näistä muodostuu hieman sipulia muistuttavia ryhmiä. Makureseptorisolut muistuttavat rakenteeltaan hivenen hansikasta. Solun yläpinnalla (ns. apikaalinen osa tai kärkiosa) kohdassa, joka ylettyy suuontelon limakalvolle, on kokoelma nukkalisäkkeitä eli mikrovilluksia. Makumolekyylien ”kaappaamiseen” erikoistuneet proteiinit, makureseptorimolekyylit, sijaitsevat mikrovillusten pinnalla. Makureseptorisolun pesäosaa kutsutaan basolateraaliseksi osaksi. Täällä sijaitsee suurin osa soluelimistä, esimerkiksi tuma (kuva 1 a – c). a) b) c) Makureseptorisoluja (mikrovillukset oikealla) makusilmussa Primaarisisia aksoneita (reseptorisolut ovat synapsiyhteydessä näihin) Kuva 1. Makureseptorisolujen sijainti ja rakenne: a) Vallinystyjä kielen takaosassa b) Yksittäinen vallinysty pitkittäisleikkauksessa (kuvassa harmaalla nystyä ympäröivä ”vallihauta”) c) Yksittäisen makusilmun hienorakenne Vallinysty kielessä Makusilmu vallinystyssä
- 2. Useimmiten makureseptorisolut ovat makujen suhteen erikoistumattomia, joten ne reagoivat yleensä moniin eri makuihin. Toisinaan kuitenkin makureseptorisolulla myös voi olla vain yksi optimimakunsa, joka synnyttää siinä impulssin. Vaihtoehtoisia optimimakuja on vain viisi eri tyyppiä. Nämä niin sanotut perusmaut ovat makea, suolainen, hapan, kitkerä ja ”umami”. Tekstin loppupuolella olevissa kuvissa (väliotsikot 1 – 5) perusmakujen aistimiseen johtavat solubiologiset mekanismit esitetään eri soluissa pelkästään selvyyden vuoksi. Makuaistimusten kulku Makureseptorisolut ovat synapsiyhteydessä aksoneihin (ns. primaariset eli ensimmäisen vaiheen aksonit), jotka kuljettavat makuviestejä kohti aivoja. Yksittäinen aksoni voi kuitenkin saada viestejä useammaltakin makureseptorisolulta. Tämä tarkoittaa sitä, että impulssien siirtyessä aksoneihin jo makureseptorisoluissa ilmenevä erikoistumattomuus voimistuu entisestään. Makujen käynnistämät hermoimpulssit ovat siis informaatiosisällöltään varsin yleisluontoisia aivoja kohti kiitäessään. Ytimenjatkeessa primaariset hermosolut yhtyvät tumakkeeseen nimeltä Gustatorinen tumake (GN). Kuhunkin GN:ssä olevaan solukeskukseen saapuu impulsseja usealta eri aksonilta. Näin ollen impulssien sisältämä makutieto hämärtyy GN:n synapseissa vielä entisestään. Jo tässä vaiheessa tuntuukin perin vaikealta ymmärtää, miten makujen tulkinta aivoissa ylipäänsä on mahdollista. GN:n jälkeen impulssit kulkevat vielä Thalamuksessa olevaan Ventral Posterior Medial –tumakkeeseen (= VPM) ja vasta täältä makuaivokuorelle. (Kuvat 2, 3 ja 4)
- 3. 1 2 Kuva 2. Alla olevassa kuvassa 3 ylempi poikkileikkaus esittää kohtaa 1, alempi kohtaa 2. Tulkitse siis alla olevaa kuvaa 3 osin tämän kuvan avulla.
- 4. Neljäs aivokammio Pyramidirata Vasen “Gustatory nucleus” Primaarisen hermosolun solukeskus Kieli Kolmas aivokammio 1. ja 2. aivokammio Vasen “Ventral Posteriol Medial nucleus” VPM Ohimolohko Päälaen- lohko Insula Primaarinen makuaivokuori Ytimenjatke Kuva 3. Makuimpulssien eteneminen kielestä aivoihin. Makuaivokuori (= Gustatory cortex) sijaitsee somatosensorisen aivokuoren alamutkassa (kuva 4) sekä insulassa (= ohimo- ja päälaenlohkon peittoon jäävä alue).
- 5. Makuimpulssien tulkinta Impulssin syntyyn ja siirtymiseen sisältyvien moninaisten juonenkäänteiden vuoksi aivoissa makujen tulkinta ei voi tapahtua suljetusti yksittäisten reseptorisolu → primaarinen aksoni → GN → VPM → yksittäinen aivosolu -linjojen perusteella. Sen sijaan tulkinta tapahtuu solu-, aksoni- ja aivosolupopulaatioiden tasolla. Asiaa voisi verrata pistekirjoitukseen, jossa pisteiden joukoilla (ei yksittäisillä pisteillä) luodaan merkityksiä. Jos jokin makureseptorisolu on erikoistunut makeuden aistimiseen, synnyttävät makeat makuaineet siinä impulsseja tiuhaan tahtiin. Jossakin toisessa solussa pelkkä makeus tuottaa impulsseja harvakseltaan, mutta kenties makea yhdessä happaman kanssa hieman tiheämmin. Makuepiteeliä kokonaisuutena voidaan ajatella pintana, josta viestejä kuljettavat aksonit muodostavat kartan isoaivojen makuaivokuorelle. Kartan eri osiin saapuvien viestien tiheyttä voidaan verrata pistekirjoituksessa käytettävien nystyröiden korkeuksiin: korkea nystyrä vastaisi tiheätä impulssisarjaa, matala nystyrä harvaa impulssisarjaa, tasainen pinta taas impulssien täydellistä puuttumista. Näin vaikkapa banaanijäätelöstä syntyy aivoihin varsin erilaiset ”makupinnanmuodot” kuin vaikkapa kanakastikkeesta. Makuaivokuoren päälle näkyvä osa Sensorinen aivokuori Motorinen aivokuori Keskiuurre Pikkuaivot Ohimolohko Kuva 4. Somatosensorinen aivokuori ja makuaivokuoren (= Gustatory cortex) sijainti sen alimmaisessa mutkassa.
- 6. Pelkkien makureseptorisolujen toiminnan lisäksi makuaistimus on luonteeltaan voimakkaan synergistinen. Tulkinnan lopputulokseen vaikuttavat ainakin ruuan rakenne (tuntoaisti), tuoksu (hajuaisti), ruuan ulkonäkö (näköaisti: ajattele vaikkapa vihreätä maitoa) sekä ruuan lämpötila (lämpöaisti). Perusmakujen (suolainen, makea, hapan, kitkerä, ”umami”) synnyttämien impulssien solubiologinen tausta Seuraavassa on tiivistelmäluontoinen esitys kunkin perusmaun taustalla olevista solubiologisista prosesseista. Useimmiten makureseptorisolut ovat makujen suhteen erikoistumattomia, joten ne voivat reagoida samanaikaisesti moniinkin eri perusmakuihin. Tämä johtuu siitä, että kussakin yksittäisessä solussa voi olla usean eri perusmaun reseptorimolekyylejä. Kuten jo aikaisemmin mainitsin, alla olevissa kuvissa (väliotsikot 1 – 5) kunkin perusmaun käynnistämät solubiologiset mekanismit esitetään tapahtuviksi eri soluissa pelkästään selvyyden vuoksi. 1. Suolainen Suolat, kuten natriumkloridi (= ruokasuola) ärsyttävät makusoluja siten, että natrium- ioneja (Na+) pääsee solun sisälle joko solun kärkiosassa (= apikaalisessa osassa) tai tyviosassa (= basolateraalisessa osassa) sijaitsevien natrium-kanavien (proteiineja) kautta. Solu depolarisoituu samalla tavoin kuin hermosolut impulssin käynnistyessä. Aivan, kuten presynaptisten hermosolujen päätenappuloissa, myös makusoluissa depolarisaatio huipentuu solujen ulkopuolella olevassa kudosnesteessä olevien kalsium- ionien (Ca+) sisäänvirtaukseen. Kyseinen tapahtuma tyypillisesti edeltää solun sisällä olevien kalvorakkuloiden yhtymistä solukelmuun (näin tapahtuu esim. allergiareaktioissa, kun syöttösolut vapauttavat ympäristöönsä histamiinia). Hermosoluissa, mutta myös makusoluissa, solukelmulle siirtyvät kalvorakkulat sisältävät hermoston välittäjäainetta. Välittäjäaine vapautuu makusolun ja hermosolun väliseen synapsirakoon. Impulssin näin lähdettyä matkaan makusilmu repolarisoi itsensä hermosoluista tutulla tavalla: kalium-kanaviksi kutsutut proteiinit avautuvat ja kalium-ioneja (K+) alkaa luikkia solusta ulos.
- 7. TUMA Mikrovillejä Ionikanava Hermosolun dendriitti Välittäjäainemolekyylejä sisältäviä kalvorakkuloita Välittäjäainemolekyylejä synapsiraossa Ruokasuola NaCl Na+ + + + Natrium- ioneja Na+ Na+ + ++ ++Kalsium- ioneja Ca++ Kalium- ioneja K+ Cl- + + 2. Hapan Hapot maistuvat happamalta, koska ne sisältävät vety-ioneja (H+). Vety-ioneja kutsutaan myös protoneiksi ja vety-ytimiksi. Nämä ionit vaikuttavat makusoluihin kolmella tavalla: ne itse siirtyvät soluihin sisälle, ne tukkivat mikrovilluksissa sijaitsevia kalium-kanavia (K+) ja ne aukaisevat muiden positiivisten ionien liikkumisen sallivia kanavaproteiineja.
- 8. Sisälle näin syöksyvien positiivisten ionien vaikutuksesta solu depolarisoituu. Lopulta myös kalsium-ioneja (Ca+) syöksyy soluun ja välittäjäainetta sisältävät kalvorakkulat tyhjentyvät synapsirakoon, kuten kohdassa yksi. TUMA Mikrovillejä Ionikanava Hermosolun dendriitti Välittäjäainemolekyylejä synapsiraossa Välittäjäainemolekyylejä sisältäviä kalvorakkuloita + + ++ + + Natrium- ioneja Na+ Kalium- ioneja K+ Kalsium- ioneja Ca++ Muita positiivisesti varautuneita ioneja Vety-ioneja H+ Vety-ionit tukkivat Kalium-kanavia H+ K+ K+K+ + + ++ 3. Makea
- 9. Makeissa ärsykkeissä (sokeri, makeutusaineet) makumolekyylit itse eivät siirry makusolujen sisälle. Sen sijaan ne kiinnittyvät solun ulkopinnalla oleviin proteiinirakenteisiin reseptoreihin. Nämä puolestaan ovat kytköksissä niin sanottuihin G- proteiineihin. Solukelmun kahden fosfolipidikerroksen välissä olevia G-proteiineja on yksittäisen reseptorin naapurustossa kolme erilaista. Makumolekyylin kiinnittyminen reseptoriinsa saa G-proteiinit aktivoitumaan. Aktiivisina ne puolestaan aktivoivat solukelmun sisäpinnalla kiinni olevan entsyymin. Tämä puolestaan aktivoi solulimassa olevia toisiolähettimolekyylejä. Toisiolähetti saa solukelmun kalium-kanavat (K+) sulkeutumaan (välillisesti). Näin solun sisälle alkaa jälleen virrata suurin määrin positiivisesti varautuneita ioneja eli solu depolarisoituu. Myös tässä kalsium-ioneja Ca+ syöksyy lopulta soluun ja välittäjäainetta sisältävät kalvorakkulat tyhjentyvät synapsirakoon.
- 10. TUMA G-proteiinivälitteinen reseptori, suurennos kuvan alareunassa Hermosolun dendriitti Välittäjäainemolekyylejä sisältäviä kalvorakkuloita Välittäjäainemolekyylejä synapsiraossa + Natrium- ioneja Na+ ++ Kalsium- ioneja Ca++ Kalium- ioneja K+ ei poistu solusta + ++ K+ Toisiolähettiä muokkaava entsyymi Toisiolähetin esimuoto Valmis toisiolähetti tukkii epäsuorasti K+-kanavia (katkonuoli ja valkoinen neliö) Sokeri- tai makeutus- ainemolekyyli Makumolekyyli reseptorissaan Alfa Beta Gamma G-proteiinikompleksi
- 11. 4. Kitkerä Kitkerät ärsykkeet, kuten kiniini, vaikuttaa sekin soluun G-proteiinien välityksellä. Tässä tapauksessa toisiolähettijärjestelmän toiminta johtaa solulimakalvoston (ER) sisällä olevien kalsium-varastojen (Ca+) tyhjentymiseen. Solulimaan muodostuu nopeasti positiivinen varaustila eli solu depolarisoituu. Ca+-ionien vaikutuksesta myös välittäjäainetta vapautuu synapsirakoon.
- 12. TUMA G-proteiinivälitteinen reseptori Hermosolun dendriitti Välittäjäainemolekyylejä sisältäviä kalvorakkuloita Välittäjäainemolekyylejä synapsiraossa + Natrium- ioneja Na+ ++ Kalsium- ioneja Ca++ Kalium- ioneja K+ + ++ Toisiolähettiä muokkaava entsyymi Toisiolähetin esimuoto Valmis toisiolähetti Kitkerä makuainemolekyyli K+ Toisiolähetti vapauttaa Ca++ - ioneja solulimakalvoston (ER) sisältä
- 13. 5. Umami (eräiden aminohappojen maku) Myös umamin maistaminen tapahtuu G-proteiinivälitteisen reseptorin avulla ja käynnistää solun sisäisen toisiolähettijärjestelmän. Toisiolähettijärjestelmän vaikutusperiaatetta ei kuitenkaan vielä tätä kirjoitettaessa tunneta. Esimerkiksi glutamaatti niminen aminohappo maistuu umamilta.
- 14. TUMA G-proteiinivälitteinen reseptori Hermosolun dendriitti Välittäjäainemolekyylejä sisältäviä kalvorakkuloita Välittäjäainemolekyylejä synapsiraossa + Natrium- ioneja Na+ ? Kalsium- ioneja Ca++ Kalium- ioneja K+ + ++ K+ Toisiolähettiä muokkaava entsyymi Toisiolähetin esimuoto Valmis toisiolähetti Glutamaatti
- 15. Maku- ja hajuaisti vertailussa Vaikka maku- ja hajuaisti ovat toistensa täydentäjiä, ovat ne geneettisessä ja solubiologisessa suhteessa varsin erilaiset. Makuaistin viittä erilaista reseptorigeeniä vastaan tuntuvat hajuaistin tuhat erilaista reseptrorigeeniä uskomattomalta tuhlaukselta. Toinen merkittävä ero on, että hajureseptorisoluista kukin ilmentää vain yhtä reseptorigeeniä kerrallaan, sen sijaan makureseptorisoluissa on yleisesti toiminnassa useitakin erilaisia reseptorigeeniversioita. Aisteissa samankaltaista on kuitenkin viestien tulkintatapa, jossa yksittäisten reseptorisolujen sijasta hyödynnetäänkin laajoja solupopulaatioita.