Lisosomas, peroxisomas, glioxisomas, ribosomas.
•Descargar como PPT, PDF•
5 recomendaciones•32,529 vistas


Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Similar a Lisosomas, peroxisomas, glioxisomas, ribosomas.
Similar a Lisosomas, peroxisomas, glioxisomas, ribosomas. (20)
Último
Último (20)
Lisosomas, peroxisomas, glioxisomas, ribosomas.
- 1. LISOSOMAS 1
- 2. 2
- 3. Lisosoma = lisis (disolución, rotura) + soma (cuerpo). Organelos presentes en todas las células eucariotas. Se distinguen de otros organelos membranosos por su forma y sus funciones Lisosomas ⇔ estabilidad en la célula viviente ⇔ membrana que rodea a las enzimas y las aisla del contenido celular. Las enzimas lisosómicas actúan en medio ácido ⇒ bomba protónica en su membrana. El pH del interior del lisosoma es de 5 La membrana es resistente a enzimas que contiene: - Todo el proceso de digestión se realiza dentro del lisosoma. - Protege al resto de la célula del efecto destructivo de los enzimas. - Su estabilidad es muy importante para el funcionamiento normal de la célula. 3
- 4. El lisosoma se autoprotege de las proteasas lisosómicas y de la acidez porque su hemimembrana interior está intensamente glucosilada. Las membranas del lisosoma tienen también receptores que le permiten reconocer las vesículas con las que se fusionan, unas proteínas LAMP (proteínas asociadas a la membrana lisosómica) y proteínas de transporte que facilitan el paso de productos finales de la degradación de sustancias hacia el citosol. 4
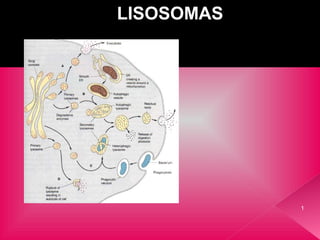
- 5. TIPOS DE LISOSOMAS Se denominan lisosomas primarios a aquellos que contienen sólo enzimas y que aún no han participado en procesos digestivos. Suelen ser muy pequeños, de 0,2-0,5 µm y corresponden a vesículas que contienen hidrolasas ácidas emanadas de la cara trans del complejo de Golgi. Los lisosomas primarios cuando se fusionan con el compartimento endosómico tardío se incluyen en la expresión de lisosomas secundarios y su contenido es heterogéneo. En la fagocitosis de bacterias u otros microorganismos por los leucocitos polimorfonucleares y macrófagos. Las vacuolas de endocitosis o fagosomas se unen a los lisosomas procedentes del compartimento endolisosomal, formándose los fagolisosomas. 5
- 6. 6
- 7. Los lisosomas también pueden digerir organelos internos de la célula como mitocondrias, membranas etc. en lo que se denominan vacuolas autofágicas, autolisosomas o citolisosomas. Esta destrucción de organelos está controlada, pero los lisosomas podrían destruir la célula entera si se rompen sus membranas, como ocurre tras la muerte (degeneración postmortem ). Los residuos de la digestión, o bien son eliminados, o se acumulan en el lisosoma y permanecen allí por el resto de la vida de la célula formando los denominados cuerpos residuales o telolisosomas. Ej. los granos de lipofuscina que se observan en células con larga vida, como las del hígado, miocardio o neuronas. 7
- 8. Las ezimas son dirigidas a los lisosomas por una señal específica. Cada enzima lisosomal contiene una señal Manosa 6-fosfato que permite seleccionarlas de la otras proteínas en la red trans Golgi y llevadas a los lisosomas por transporte vesicular. La enzima cis-Golgi NAcGlc fosfotransferasa primero añade un grupo NAc Glc-fosfato al sexto carbono de un residuo de manosa en la cadena oligosacárida. La NAcGlc es luego removido dejando un fosfato en la manosa. 8
- 9. FUNCIONES Digestión de materiales que provienen del ambiente extracelular. Algunos parásitos intracelulares y microbios patogénicos han adaptado la via heterofágica para su propia ventaja: “Escapar”: Secreción de toxinas que deterioran la membrana lisosomal (Shigella flexneri, Listeria monocytogenes, Ricketsia, ricketssi). “Evadir”: Inhibición de fusión fagosoma-lisosoma (Salmonella typhimurium, Mycobacterium tuberculosis. Inhibición de acidificación de fagolisosoma (especies de Mycobacterium). La cloroquina, que es un antipalúdico, incrementa el pH interno del lisosoma inactivando sus enzimas. El dextrano y polivinilo (sustitutos del plasma) y el sílice (exposición ambiental) se acumulan al no existir enzimas para su degradación. La vitamina A es labilizante (rompe la membrana), mientras que 9 la
- 10. Renovación de células y del material extracelular. Intervienen en la crinofagia. Son importantes en las células germinales y en fertilización. En la síntesis de hormonas. 10
- 11. Los peroxisomas, morfológicamente son parecidos a los lisosomas: es decir, partículas esféricas limitadas por una membrana Con un diámetro variable entre 0.3 y 1.5 μm de diámetro, y con un contenido enzimático. En el interior de algunos pueden observarse estructuras cristalinas que generalmente corresponden a la enzima urato oxidasa. La membrana del peroxisoma posee transportadores de electrones como el citocromo b5, y las enzimas NADH-citocromo b5 reductasa y NADH-citocromo P450 reductasa. Su composición es similar a la del retículo endoplasmático. Presentan una matriz formada por proteínas enzimáticas, muchas de ellas peroxidasas y entre las que nunca suelen faltar la catalasa 11
- 12. Contienen enzimas que utilizan el oxígeno molecular para eliminar átomos de hidrógeno de sustratos específicos, a través de una reacción oxidativa que produce H2O2 R + O2 R + H2O2 El H2O2 resultado de la reacción es un producto altamente tóxico que es eliminado por otra enzima del peroxisoma, la catalasa, según la reacción: 2 H2O2 2 H2O + O2 FUNCION Actividad enzimática Catabolismo de las purinas β-oxidación de los ácidos grasos Detoxificación 12
- 13. Se cree que se originarían como una gemación de RER desprovista de ribosomas. En la membrana del peroxisoma hay unas proteínas que son comunes a la membrana del RER y a la del peroxisoma. También se cree que sean capaces de reproducirse, previo crecimiento seguido de fisión. En tal caso los componentes de la membrana serían importados del citoplasma a través de proteínas translocadoras. 13
- 14. Reacciones que tienen lugar en el peroxisoma Reacciones catabólicas Respiración celular basada en el peróxido de hidrógeno Catabolismo de las poliaminas Catabolismo de las purinas Oxidación del etanol Oxidación del ácido L-pipecólico * Beta-oxidación Ácidos grasos de cadena de más de 8 carbonos Ácidos grasos de cadena muy larga * Ácidos dicarboxílicos de cadena larga Prostaglandinas Xenobióticos Cadena lateral del colesterol Alfa-oxidación Ácido fitánico * Ácido pristánico * Reacciones anabólicas Biosíntesis de los plasmalógenos * Biosíntesis del colesterol Biosíntesis de los ácidos biliares * Gluconeogénesis Transaminación del glioxalato * * Indica que ciertos pasos de la vía metabólica tienen lugar exclusivamente en el peroxisoma. 14
- 15. La presencia de catalasa y peroxidasa son las que usan los peroxisomas en el hígado para descomponer las moléculas de alcohol en sustancias que puedan ser eliminadas del organismo. Aproximadamente una cuarta parte del alcohol que entra en el hígado se procesa en los peroxisomas. Todas las proteínas peroxisomales se sintetizan en polirribosomas libres, entran en el citosol y contienen peptido señal de entrada peroxisomal (SEP, o PTS del inglés Peroxisomal Targeting Signals) que los dirigen hacia el organelo. Ciertas deficiencias involucran a peroxisomas ineficaces, tales como la enfermedad de Zellweger. Es una enfermedad extremadamente rara que pertenece al grupo de las enfermedades peroxisomales y se caracteriza por asociar alteraciones neurológicas graves con dimorfismo (forma defectuosa de un aparato u órgano) craneofacial debido a una ausencia de peroxisomas por una anomalía de su biogénesis en las células renales, hepáticas y fibroblastos 15
- 16. Son vesículas típicas de los vegetales. Están presentes en las semillas donde actúan durante la germinación. Sus enzimas convierten lípidos a glúcidos y de estos, se obtiene energía. Las reacciones químicas de estos procesos son conocidas como el ciclo de glioxilato. 16
- 17. CICLO DEL GLIOXILATO El ciclo del glioxilato es una variante del ciclo del ácido cítrico (concretamente un "by-pass" de las estapas descarboxilantes) que ocurre en losglioxisomas de las células vegetales (también ocurre en muchos hongos y protozoos). Permite generar glucosa a partir de ácidos grasos , esto es muy importante en las semillas, debido a que la mayor parte de la energía metabólica necesaria para su desarrollo se encuentra en forma de triacilgliceroles. 17
- 18. REACCIONES BIOQUÍMICAS 1) La acetil-CoA (procedente de la oxidación de ácidos grasos) reacciona con el oxalacetatoformando citrato. La enzima que cataliza esta reacción es la citrato sintasa*. 2) El citrato reacciona con la enzima aconitasa* formando Isocitrato. 3) El isocitrato, mediante una reacción catalizada por la enzima isocitrato liasa, se fragmenta en glioxilato y succinato. 4a) El succinato es metabolizado en forma similar que en el ciclo del ácido cítrico a fumarato mediante la enzima succinato deshidrogenasa y luego a malato por la enzima fumarasa. 4b) La acetil-CoA transfiere un acetilo al glioxilato produciendo malato en una reacción catalizada por la enzima malato sintasa*. 5) El malato se deshidrogena para formar nuevamente oxalacetato mediante una reacción catalizada por la enzima malato deshidrogenasa. El oxalacetato es capaz de generar glucosa mediante gluconeogénesis. 18
- 19. Depósitos celulares de agua, sales y solutos. En los vegetales suelen fusionarse y formar una gran vacuola que ocupa casi todo el citoplasma. Su membrana se denomina tonoplasto. 19
- 20. Contenido vacuolar: El contenido de las vacuolas es muy variable. Depende de la planta, de la célula (dentro de la planta) y del estado fisiológico de la célula. A demás hay compuestos que se almacenan de forma permanente en la vacuola y otros que se intercambian periódicamente con el citoplasma. Se puede encontrar iones (K , Mg , Ca , Cl ), también ácido orgánicos, proteínas, mucílagos, pigmentos (antocianinas), 20 heterósidos.
- 21. Lugar de almacén de sustancias dañinas (ejm: ácido sulfúrico) que las célula no puede excretar. Lugar de almacén de azúcares y aminoácidos que la célula no necesita de manera inmediata Almacena pigmentos azul y púrpura dando color a muchas flores. 21
- 22. 22
- 23. Existen dos tipos de vacuolas: › Vacuola pulsátil o de expulsión: regula la concentración de agua en la célula. › Vacuola alimenticia: digestión celular. 23
- 24. Están constituidos por dos subunidades: mayor (60S) y menor (40S) en conjunto 80S. Se sintetizan en el nucléolo. 24
- 25. Están compuestos por ARN ribosómico y proteínas. Se encuentran: › Asociadas con el RER › En la superficie externa de la carioteca › Dispersos en el citoplasma › Dentro de las mitocondrias y cloroplastos. 25
- 26. Al conjunto de ribosomas en el citoplasma unidos a un ARNm se le denomina polisomas. 26
- 27. FUNCIÓN: Realizan el proceso de la traducción en la síntesis de proteínas. 27
- 28. Selección de Mercancía, proteínas de revestimiento y gemación Formación y Fusión de una vesícula de Transporte 28
- 29. Selección de Mercancía, proteínas de revestimiento y gemación Proteínas de unión a GTP: Important es para la formació n de vesículas 29
- 30. Vesículas Revestidas: Funciones 3 Tipos Vesículas revestidas Vesículas revestidas por Clatrina de COP II Vesículas revestidas de COP I 30
- 31. Incorporación de proteínas lisosómicas a vesículas revestidas 31
- 32. Iniciación de una vesícula revestida por Clatrina 32
- 33. Fusión de las Vesículas 1ro. Reconocimiento correcto de la vesícula a su membrana diana. 2do. Fusión de La membrana de la vesícula y la membrana diana deben fusionarse. (proteínas: v-SNARE y t-SNARE) Proteínas Rab 33
- 34. Fusión de Vesículas 34
- 35. 35
- 36. 36
Notas del editor
- Mecanismo de transporte de las vesículas El transporte de las vesículas es una actividad celular fundamental, responsable del tráfico molecular entre diversos compartimientos rodeados de membrana. La selectividad es clave para mantener la organización funcional de la célula. Son necesarios tres procesos en cada paso de la vía secretora. Las proteínas mercancía son separadas de las proteínas dirigidas a otros destinos. Una yema que contiene la mercancía debe desarrollarse en la membrana y separarse de ella Esa vesícula debe moverse hacia la membrana diana y fusionarse con ella
- La formación de la mayoría de las vesículas de transporte está regulada por proteínas de unión a GTP, a través de proteínas adaptadoras que interaccionan directamentecon una proteína de revestimiento vesicular. Entre estas proteínas de unión a GTP están ARF1, Sar1 y la gran familia de proteínas Rab.
- Vesículas de revestimiento de clatrina: son responsables de la internalización de moleculas extracelulares por endocitosis, además del transporte de moléculas desde la red del trans Golgi hacia los endosomas y lisosomas. COP indica proteína de revestimiento “coat protein” COPI funcionan en los mecanismos de recuperación que retienen a las proteínas residentes del Golgi y el RE COPII se forma a partir del RE y transporta su mercancía a lo largo de la vía secretora hasta el aparato de Golgi.
- Las proteínas dirigidas a los lisosomas están marcadas por manosa-6-fosfato en la red trans de Golgi. Los receptores de manosa-6-fosfato atraviesan la membrana del Golgi y actúan como sitios de unión para las proteínas adaptadoras citosólicas, que a su vez se unen a la clatrina. Las clatrinas están constituídas por tres cadenas proteicas que se asocian entre sí para formar una red semejante a la de las canastas de baloncesto, que distorsiona la membrana y dirige la gemación de las vesículas.
- La proteína pequeña de unión a GTP, ARF, puede iniciar la formación de una vesícula revestida de clatrina en la membrana del trans de Golgi. Una vez transportada a la membrana, ARF/GDP es activada por un factor de intercambio de nucleótidos de guanina. ARF/GTP recluta a la membrana a una proteína adaptadora GGA, y esta proteína recluta al receptor de manosa-6-fosfato, que transporta a la hidrolasa lisosómica, interaccionando con la cola citoplásmica del receptor. GGA también recluta a una segunda proteína adaptadora AP1, que sirve como sitio de unión para el ensamblaje de una cubierta de clatrina sobre la vesícula.
- La localización correcta de las proteínas Rab es clave para establecer la especificidad del transporte.
- La fusión vesicular es iniciada por Rab/GTP. Rab/GTP específicas presentes en la membrana vesicular y en la membrana diana unen proteínas efectoras para anclar la vesícula a la membrana diana. El anclaje permite que interaccionen las vSNAREs y las tSNAREs, lo que proporciona la energía necesaria para aproximar las membranas. Esta proximidad de las membranas desestabiliza las bicapas lipídicas y la vesícula y la membrana diana se fusionan. Cambios en las interacciones proteína-proteína reclutan a NSF y las SNAPS al complejo SNARE, y desensamblan el complejo empleando energía obtenida de la hidrólisis de ATP.