

Recomendados
Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Más de Pasi Vilpas
Más de Pasi Vilpas (20)
Hermosolun toiminta
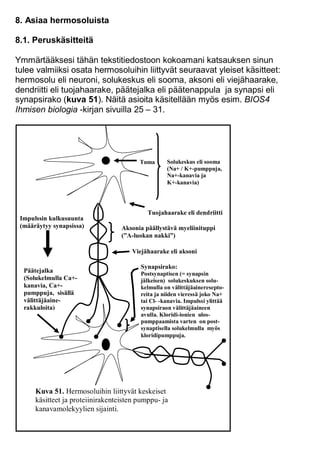
- 1. 8. Asiaa hermosoluista 8.1. Peruskäsitteitä Ymmärtääksesi tähän tekstitiedostoon kokoamani katsauksen sinun tulee valmiiksi osata hermosoluihin liittyvät seuraavat yleiset käsitteet: hermosolu eli neuroni, solukeskus eli sooma, aksoni eli viejähaarake, dendriitti eli tuojahaarake, päätejalka eli päätenappula ja synapsi eli synapsirako (kuva 51). Näitä asioita käsitellään myös esim. BIOS4 Ihmisen biologia -kirjan sivuilla 25 – 31. Tuojahaarake eli dendriitti Viejähaarake eli aksoni Päätejalka (Solukelmulla Ca+- kanavia, Ca+- pumppuja, sisällä välittäjäaine- rakkuloita) Tuma Solukeskus eli sooma (Na+ / K+-pumppuja, Na+-kanavia ja K+-kanavia) Synapsirako: Postsynaptisen (= synapsin jälkeisen) solukeskuksen solu- kelmulla on välittäjäaineresepto- reita ja niiden vieressä joko Na+ tai Cl- -kanavia. Impulssi ylittää synapsiraon välittäjäaineen avulla. Kloridi-ionien ulos- pumppaamista varten on post- synaptisella solukelmulla myös kloridipumppuja. Impulssin kulkusuunta (määräytyy synapsissa) Kuva 51. Hermosoluihin liittyvät keskeiset käsitteet ja proteiinirakenteisten pumppu- ja kanavamolekyylien sijainti. Aksonia päällystävä myeliinituppi (”A-luokan nakki”)
- 2. Hermosolut ovat monella tavalla erikoistuneita. Niiden ainutlaatuisin piirre on kyky synnyttää solukalvollaan sähköisiä muutoksia (jännitteen muutoksia) ja lisäksi kyky siirtää muutos naapurihermosoluihin, joskus myös muihin solutyyppeihin. Hermosolun solukelmu työskentelee ankarasti. Aikaisemmilta biologian kursseilta muistat varmaankin, että solun (ja siis elämän) työkalut ovat proteiineja. Varsinaisesti työn siis solukelmussa tekevät kelmuun kiinnittyneet proteiinit. Geeni puolestaan on sellainen jakso DNA-rihmaa, joka sisältää yhden proteiinin rakenneohjeen. Siksi myös hermosolujen toiminnalla on geneettinen perusta. Hermosoluille ominaisten proteiinien tekemä työ johtaa siihen, että solukelmun ulkopuolelle muodostuu positiivinen varaus suhteessa sisäpuolella vallitsevaan negatiiviseen varaustilaan. Varausero on nimeltään lepojännite. Kaikkialla solukelmussa on myös proteiineja, jotka tietyssä tilanteessa sallivat varausten vaihtaa paikkaa. Tuolloin jännitteen suunta muuttuu. Solukelmua pitkin eteneviä varaustilan muutoksia kutsutaan hermoimpulsseiksi. Kahden hermosolun välissä olevan synapsiraon kohdalla solukelmun osat lorauttelevat ulkopuolelleen välittäjäaineita, toiset solukelmun osat taas vastaanottavat niitä ja reagoivat niihin jännitteen muutoksella. Jännite syntyy aina positiivisten ja negatiivisten sähkövarausten välille. Jännitteen eli potentiaalin arvo ilmoitetaan voltteina (V). Soluihin, tarkasti ottaen solukelmun ulko- ja sisäpuolen välille, jännite syntyy, kun sisä- ja ulkopuolella (=kudosnesteessä) on eri määrät ioneja tai jos solukelmun eri puolilla olevien ionien sähkövaraus on vastakkaismerkkinen. Hermosolun kannalta tärkeimmät ionit ovat: - Na+ ioni, runsaasti kuosnesteessä, vähän solulimassa - K+ ioni, runsaasti solulimassa, vähän kudosnesteessä - Cl- ioni, runsaasti kudosnesteessä, vähän solulimassa - Ca+ ioni, jonkin verran kudosnesteessä, vähän sisäpuolella
- 3. - negatiivisesti varautuneet makromolekyylit (=proteiinit), runsaasti solulimassa, hyvin vähän kudosnesteessä 8.2. Lepojännite (kuva 52) Kun kaikki varaukset lasketaan yhteen, voidaan sanoa, että hermosolun sisäpuoli on negatiivisesti, ulkopuoli positiivisesti varautunut. Tällaisessa solussa solukalvon yli vallitsee ns. lepojännite, jonka suuruus on -70mV (jännite mitataan sisäpuolelta siksi se on negatiivinen). Itse asiassa kaikissa soluissa vallitsee jonkinlainen lepojännite, mutta hermosoluissa se on erityisen suuri. Ja vain hermosoluissa jännite voi muuttua salamannopeasti johonkin toiseen arvoon. Tällöin sanotaan, että hermosolussa ilmenee hermoimpulssi. Ionipitoisuuksien erot eivät synny itsestään, vaan solukelmussa olevat proteiinirakenteiset pumput siirtävät varsinkin Na+ ja K+ ioneja eri suuntiin. Lisäksi (useimmiten negatiivisesti varautuneita) makromolekyylejä esiintyy enimmäkseen solujen sisäpuolella, jossa ne valmistetaan. Ionien siirtotyö kuluttaa runsaasti ATP:tä. Lepotilassa jopa 25% elimistön ATP-kulutuksesta menee tähän puuhaan.
- 4. Lepojännite siis perustuu solukelmun kykyyn pumpata ioneja ja toisaalta estää niiden passiivinen siirtyminen kelmun läpi takaisin. Ionit todella pysyvät paikoillaan, vaikka niiden pitoisuuserot ovat hyvinkin suuria. 8.3. Yhteenveto hermosolun solukelmun molekyyleistä (Kaikki muut ovat proteiineja paitsi kohta 1. Fosfolipidi-kolesterolikalvo): 1. Fosfolipidi-kolesterolikalvo - solun ulkokalvo eli solukelmu, joka toimii sähköeristimenä, koska ei läpäise ioneja lainkaan Natrium / Kalium –pumppu - pumppaa kolme Na+-ionia ulos samalla kun vain kaksi K+-ionia sisään → solun ulkopuoli muuttuu varaukseltaan positiiviseksi solun sisäpuoleen verrattuna (= lepojännite) K+ -kanava - K+-kanavat ovat kiinni lepojännitteen aikana Na+-kanava - Na+-kanavat ovat kiinni lepojännitteen aikana Hermosolun sisäpuoli Hermosolun ulkopuoli Solukelmu ATP ADP + P K+ K+ Na+ Na+Na+ K+ K+ K+ K+ K+ K+ Na+ Na+ Na+ Na+ Na+ Na+ Kuva 52. Hermosolun toiminta lepojännitteen aikana.
- 5. 2. Natrium-kalium-pumppu eli Na/K-ATP-aasi - pumppaa kerralla kaksi K+ionia sisään soluun, kolme Na+ionia ulos ja käyttää tähän työhön yhden ATP-molekyylin, joka hajoaa ADP:ksi ja fosfaatiksi - näitä esiintyy koko solukelmun alueella, yhteenlaskettu määrä lienee n. miljoona / hermosolu, tehtävänä lepopotentiaalin ylläpito 3. Na-kanava - avautuvat jännitteen hiemankin muuttuessa (välittäjäaineen vaikutuksesta tai joskus jopa solun sisäisten kemiallisten muutosten takia) ja sallivat Na+ionien virrata niiden pitoisuus- ja/tai varauseron määräämään suuntaan 4. K+kanava - kuten Na+kanavat 5. Ca+kanava - kuten Na+ ja K+kanavat, mutta näitä esiintyy aksonin päätejaloissa synapsirakojen alueella 6. Cl-kanava - kuten Na+, K+ tai Ca+kanavat, mutta näitä esiintyy vain dendriittien ja sooman eli solukeskuksen alueella välittäjäainereseptorien naapureina 7. Välittäjäainereseptori - näitä on useita eri tyyppejä, esiintyvät dendriittien ja sooman solukelmulla , näiden naapurina esiintyy usein joko Na+ tai Cl- kanava - välittäjäainereseptoreilla voi toisinaan olla myös toisiolähettivaikutus 8. Ca+ pumppu ja Cl-pumppu - sekä Ca+ että Cl-ionien aktiiviseen liikuttelemiseen solukelmulla on ATP-käyttöisiä pumppuproteiineja, näitä on solukelmussa erityisesti synapsirakojen alueella 8.4. Ionien liike solukelmun läpi
- 6. Kun hermosolussa, jossa vallitsee lepojännite, aukeaa ionikanavia, ionit alkavat siirtyä solukalvon puolelta toiselle kahden erilaisen voiman yhteisvaikutuksesta. Nämä voimat ovat a) Coulombin voima: positiiviset ionit pyrkivät kohti negatiivista varausta ja päinvastoin b) pitoisuuserosta aiheutuva voima: ioni siirtyy sieltä, missä sitä on paljon sinne, missä sitä on vähän Huomaa, että useissa tapauksissa voimat vaikuttavat eri suuntiin. Kun näin on, ionit ratkaisevat tilanteen hakeutumalla sellaisiin asemiin, joissa vastakkaisiin suuntiin vaikuttavat voimat ovat yhtä suuret. Lepäävässä hermosolussa K+ioni on tällaisessa asemassa: vaikka lepojänniteen aikana osa K+kanavista onkin auki K+ionit pysyvät solussa siitä huolimatta, että pitoisuuseron luulisi ajavan niitä solusta ulos. Selityksenä on, että sisäpuolella vallitsee negatiivinen kokonaisvaraus ja se vetää K+ioneja puoleensa. Muut ionit eivät tuolloin (siis lepojännitteen vallitessa) ole tasapainossa, mutta nepä eivät pääsekään liikkumaan, koska kanavat ovat kiinni. 8.5. Toimintajännite eli impulssi eli miten lepojännite purkautuu ? Kun hermosolua ärsytetään, siinä saattaa käynnistyä impulssi. Impulssiksi kutsuttu jännitemuutos tapahtuu useassa, osin samanaikaisesti tapahtuvassa vaiheessa. Nämä ovat seuraavat: 1. Na+kanavia aukeaa paikallisesti, yleensä aksonin tyvessä. Tällöin Na+ioneja alkaa virrata solun sisään, koska sekä Coulombin voima, että pitoisuusero vaikuttavat samaan suuntaan. 2. Na+ioneja siirtyy niin paljon, että solun sisäpuoli muuttuu varaukseltaan positiiviseksi ja ulkopuoli negatiiviseksi (koska Na+ionit ovat poistuneet sieltä). Nyt Na+ionipitoisuus on tasapainossa siihen vaikuttavien voimien suhteen. Solun sisäpuolelta mitattuna jännitteen suuruus on nyt +35 mV ja solussa vallitsee toimintajännite eli hermoimpulssi tällä solukelmun kohdalla. 3a. Jänniteherkät K+kanavat avautuvat ja K+ioneja alkaa siirtyä solun ulkopuolelle pitoisuuseron ja ulkopuolella nyt vallitsevan negatiivisen varauksen ajamina. Tämä jatkuu, kunnes K+ionipitoisuus on uudessa tasapainotilassa ja lepojännite on palautunut solukelmun tälle kohdalle.
- 7. 3b. Samaan aikaan ensimmäisinä avautuneiden Na+kanavien lähistöllä olevat Na+kanavat aukeavat jännitemuutoksen ärsyttäminä. Näin hermoimpulssi alkaa edetä pitkin aksonia. K+ -kanavat - K+-kanavat avautuvat toimintajännitteen jälkimmäisen vaiheen aikana Na+-kanavat - Na+-kanavat avautuvat toimintajännitteen alkaessa Hermosolun sisäpuoli Hermosolun ulkopuoli Solukelmu K+ K+ Na+ Na+Na+ K+ K+ K+ K+ K+ K+ Na+ Na+ Na+ Na+ Na+ Na+ Kuva 53. Hermosolun toiminta toimintajännitteen aikana. Toimintajännitteen päätyttyä aloittaa Na+ / K+ -pumppu jälleen toimintansa niin, että hermosolu saavuttaa uudelleen lepojännite- tilanteen Aksonien tehtävänä on kuljettaa impulssi hermosolun loppuun synapsiin asti. Aksonit toimivatkin ”kaikki tai ei mitään” –periaatteella. Kun niissä tapahtuu jännitemuutos, se on aina samansuuruinen eli n. +35 mV. Hermosolun muissa osissa, solukeskuksessa ja dendriiteissä lepojännite on samansuuruinen kuin aksonissa, mutta tilanteesta riippuen tämä jännite voi muuttua negatiivisemmaksi tai positiivisemmaksi. Negatiiviseksi muuttuminen tarkoittaa sitä, että lepojännite voimistuu. Mitä positiivisemmaksi jännite muuttuu, sitä todennäköisemmin hermosolussa syntyy impulssi. 8.6. Synapsi Jännitteen muutokset kumpaankin edellä mainittuun suuntaan tapahtuvat useimmiten synapsivaikutuksen (siis muista hermosoluista
- 8. saapuvien välittäjäaineiden) tuloksena. Kun impulssi etenee synapsirakoon asti, ratkaistaan sen eteneminen tai pysähtyminen uudelleen seuraavan hermosolun pinnalla olevien satojen tai tuhansien synapsien yhteisvaikutuksen perusteella. Tätä tarkoittaa sanonta ”hermosolu toimii laskukoneena” (kuva 54). Hermosolujen toimintaa käsittelevässä kirjallisuudessa esiintyy usein käsitepari pre- ja postsynaptinen hermosolu (=neuroni). Synapsissa presynaptinen (=synapsia edeltävä) hermosolu vaikuttaa välittäjäainemolekyylien välityksellä postsynaptiseen (= synapsin jälkeiseen) hermosoluun. Synapseja on tavallisimmin aksonin ja dendriitin välillä, mutta niitä voi olla myös aksonin ja sooman tai jopa kahden aksonin välillä. Synapsin kaltaisia ovat myös seuraavat liittymät (impulssin etenee ensin mainitusta solutyypistä jälkimmäiseen): - hermosolu – poikkijuovainen lihassolu - hermosolu – sileä lihassolu - hermosolu – rauhassolu - aistireseptorisolu – hermosolu 8.7. Mitä synapsin alueella tapahtuu? Myeliinituppi Kuva 54. Pieni osa yksittäiseen hermosoluun saapuvista synapsiyhteyksistä.
- 9. Synapsissa tapahtuu seuraavia asioita (vaiheet 1 – 6): 1. Toimintajännite eli aktiopotentiaali saapuu päätejalan solukelmulle. Päätejalan solukelmulla sijaitsevat jännitemuutoksille herkät Ca+-kanavat avautuvat. 2. Ca+-ioneja virtaa kudosnesteestä solun sisäpuolelle (vaihe 1) + + + + _ _ __ + _ + _ _ + _ + _ Ca+ Ca+ Ca+ Postsynapti nen neuroni Presynap- tinen neuroni Impulssin eli toiminta- jännit- teen etenemis- kohta Vaihe 1. Impulssin saapuminen presynaptisen neuronin päätejalan alueelle. Päätejalan alueella solun sisälle syöksyy natriumin sijasta kalsium-ioneja. Välittäjäaine- reseptori Mitokondrio 3. Ca+-onien sisääntulo saa välittäjäainetta sisältävät kalvorakkulat yhtymään solukelmuun, jolloin välittäjäaineet eksosytoidaan eli ”oksennetaan” synapsirakoon (vaihe 2).
- 10. Asetylikoliini-molekyylejä vapautuu synapsirakoon Postsynap- tinen neuroni Presynaptinen neuroni Vaihe 2. Impulssin eteneminen synapsiraon kohdalla. Presynaptisen hermosolun sisälle syöksyvä kalsium saa välittäjäainetta sisältävät kalvorakkulat yhtymään solukelmuun ja purkamaan välittäjäainemolekyylinsä synapsirakoon. Välittäjäaine- reseptori Mitokondrio 4. Välittäjäaineesta ja reseptorien rakenteesta riippuen tapahtuu jokin seuraavista vaihtoehdoista: a) reseptorien yhteydessä olevat Na+kanavat avautuvat eli aktiopotentiaali alkaa levitä myös postsynaptisen hermosolun solukelmulla (vaihe 3) b) reseptorien yhteydessä onkin Cl-kanavia, jolloin Cl-ionit ryntäävät postsynaptisen hermosolun sisälle eli lepopotentiaali voimistuu (Cl-ionien negatiivinen varaus muuttaa soluliman varaustilan entistä negatiivisemmaksi, siis lepopotentiaalin suuntaan), tässä on kyseessä estävä eli inhiboiva synapsi
- 11. c) reseptoreihin liittyvät kalvoproteiinit synnyttävät postsynaptisessa hermosolussa toisiolähettiä, esim cAMP:tä, joka voi aiheuttaa siihen pitempiaikaisia vaikutuksia Koska jokaisella hermosolulla on lukuisia synapsikytkentöjä muihin hermosoluihin, ei yhden synapsin toiminta riitä aiheuttamaan dendriittien tai sooman depolarisaatiota eli synnyttämään impulssia (edellä ollut kuva 54). Asetylikoliini-molekyylit sitoutuvat reseptoreihinsa, impulssin eteneminen käynnistyy postsynaptisen neuronin solukelmulla Postsynap- tinen neuroni Presynaptinen neuroni Vaihe 3. Impulssin saapuminen postsynaptisen neuronin tuojahaarakkeen tai solukeskuksen alueelle. Tyhjiä välittäjä-aine- rakkuloita Na+ Na+ Na+ + + ++ + + _ _ _ _
- 12. Postsynap- tinen neuroni Presynaptinen neuroni Vaihe 4. Impulssi alkaa edetä postsynaptisen neuronin solukelmulla. Mitokondrio Tyhjiä välittäjä- ainerakkuloita + + + + _ _ _ _ Impulssin eli toimintajännit- teen etenemis- kohta
- 13. Postsynap- tinen neuroni Presynaptinen neuroni Vaihe 5. Asetylikoliiniesteraasi hajottaa välittäjäainemolekyylit koliiniksi ja etikkahapoksi (etikkahappo + koliini = asetylikoliini). Tässä muodossa välittäjäainemolekyylit imetään takaisin presynaptisen neuronin sisälle. Mitokondrio Tyhjiä välittäjä- ainerakkuloita Etikkahappo Koliini
- 14. 8. 8. Välittäjäaineet Hermosolujen valmistamia välittäjäaineita on pitkään tunnettu kymmenittäin. Ne muistuttavat vesiliukoisia (siis aminohappo- tai proteiinirakenteisia) hormoneita rakenteeltaan ja toiminnaltaan. Postsynap- tinen neuroni Presynaptinen neuroni Vaihe 6. Koliini- ja etikkahappomolekyyleistä valmistetaan uutta välittäjäainetta. Tapahtuma kuluttaa ATP:tä, jota mitokondriot tuottavat soluhengityksen avulla. Välittäjäainerakkulat jäävät odottamaan uuden impulssin saapumista. Mitokondrio ATP
- 15. Noradrenaliini, adrenaliini, dopamiini ja serotoniini ovat amiineja (tarkkaan ottaen tyrosiini-nimisen aminohapon muunnelmia), glutamaatti, gamma-aminovoihappo (GABA) ja glysiini ovat aminohappoja. Yksi yleisimmistä välittäjäaineista on etikkahappojohdannainen nimeltä asetylikoliini. Kukin hermosolu valmistaa useita välittäjäainetyyppejä. Yksittäinen päätejalkakin saattaa tuottaa useata eri välittäjäainetta, mutta postsynaptiseen hermosoluun kohdistuvalta vaikutustavaltaan ne kuitenkin tuolloin ovat vain joko ekshiboivia tai inhiboivia. Samallekin välittäjäaineelle voi eri synapseissa olla eri tavoin vaikuttavia reseptoreita. Esimerkiksi lihassoluissa on asetylikoliinille reseptoreita, joihin liittyy Na+kanavia, mutta muissa soluissa reseptoreihin liittyykin G-proteiini- ja toisiolähettijärjestelmä, jolloin asetylikoliinin vaikutukset ovat hitaita, monipuolisia ja pitkäkestoisia. Viime aikoina on havaittu, että hermosolut viestivät toisilleen myös muiden kuin tavanomaisten välittäjäaineiden välityksellä. Tällaisia erikoisemman puoleisia viestiaineita ovat mm. endorfiinit ja ACTH (=adrenokortikotrooppinen hormoni). Nämä ovat neuropeptidejä eli isoista proteiineista leikattuja pikkuproteiineja. Ne kulkeutuvat aksonin päätteisiin kalvorakkuloissa. Leikkaaminen, sivumennen sanoen, tapahtuu Golgin laitteessa, hii! Neuropeptidien reseptoreihin ei liity ionikanavia, eivätkä ne siis voi aiheuttaa nopeita jännitemuutoksia kohdesoluissaan. Sen sijaan neuropeptidit synnyttävät toisiolähettejä (G-proteiinijärjestelmän välityksellä) tai saavat aikaan kohdeproteiiniensa fosforyloitumisen (eli aktivoitumisen). Kummassakin tapauksessa vastaanottavan solun yleinen toiminta muuttuu. Välittäjäaineiden synteesi tapahtuu hermosolun solukeskuksessa valmistuvien proteiinien avulla. Nämä työkalut puolestaan voivat siirtyä minne tahansa koko hermosolun alueella ja valmistaa siellä asianomaisia välittäjäaineita. Ennen niiden varsinaista käyttöä synapsissa välittäjäaineet on kuitenkin ensin pakattava kalvorakkuloihin, joita saadaan Golgin laitteesta tai ER:stä. Näissä kalvorakenteisissa soluelimissä on proteiineja, jotka saavat niistä kuroutumaan irti pieniä kalvorakkuloita eli vesikkeleitä. Välittäjäainemolekyylit päätyvät kalvorakkuloiden sisälle.
- 16. Toiset proteiinit (VAMPit ja Syntaxinit) avustavat kalvorakkulan sulautumisessa solukelmuun. Tällöin välittäjäainemolekyylit purkautuvat synapsirakoon. BI5-kurssin työalueellamme on luentovideo otsikolla ”Kalvorakkulat liikkeessä”. Siinä rER:in ja Golgin laitteen toiminta esitellään kattavasti. Koska jo yhdessä rakkulassa on tuhansia välittäjäainemolekyylejä, solut pyrkivät kierrättämään niitä. Aksonin päätejalka pystyy imemään (solukelmulla olevien pumppuproteiinien avulla) synapsiraosta välittäjäaineita, jotka pakataan sitten tyhjiin kalvopusseihin. Näitä saadaan päätenappulan alueella esim. solukelmusta kuromalla. Välittäjäaineet Hermoston välittäjäaineet jaetaan kolmeen ryhmään. Vaikka näistä vain viimeisen ryhmän edustajat ovat proteiinirakenteisia, on hyvä pitää mielessä, että kaikkia muitakin välittäjäaineita valmistetaan proteiinien avulla. Ja proteiinien rakenneohjeethan ovat geeneinä solun tumassa. 1. Aminohapot Gamma-amino-butyyrihappo (GABA) Glutamaatti (Glu) Glysiini (Gly) 2. Amiinit eli aminohappomuunnelmat Asetylikoliini (ACh) - asialla kaikissa lihassynapseissa Dopamiini (DA) Epinefriini eli adrenaliini Histamiini Norepinefriini (NE) Serotoniini (5-HT) 3. Peptidit Kolekystokiniini eli Cholecystokinin (CCK) Dynorphin Enkefaliinit (Enk) N-Asetylaspartylglutamaatti (NAAG) Neuropeptidi Y Somatostatiini Substance B (siis suomeksi aine B)
- 17. Thyreotropiinia vapauttava hormoni Suoliston vasoaktiivinen polypeptidi (VIP) Sellainen pläjäys oli siis katsauksemme hermosolun toimintaan! .