El documento presenta una discusión sobre las hipótesis del origen de la agricultura. Señala que la domesticación de plantas fue un proceso coevolutivo que precedió a la agricultura. Luego describe tres etapas de la domesticación: primigenia, locacional y preagrícola. Finalmente realiza un recuento de las principales hipótesis propuestas desde el siglo XIX para explicar la transición de la caza y recolección a la agricultura, destacando las ideas de Lamarck, Darwin y Childe, y la necesidad de un enfoque






























![32 Arqueo·Ciencias
Variable Ecuación Estadísticos complementarios
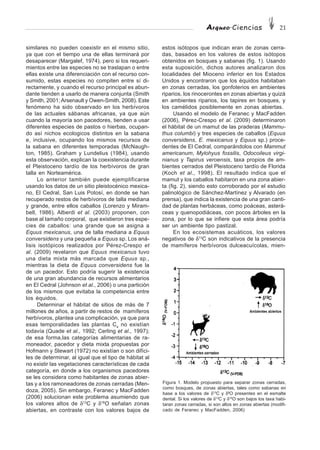
I. Alzada (o altura a la cruz, H)
I.1 Koudelka (1885, in Harcourt, 1974). Ecuación y= bx
Húmero H (mm)= 3.37·LT (mm)
Radio H (mm)= 3.32·LT (mm)
Ulna H (mm)= 2.67·LT (mm)
Fémur H (mm)= 3.01·LT (mm)
Tibia H (mm)= 2.92·LT (mm)
I.2 Harcourt (1974) (n=34) (valores de r entre 0.98 y 0.99). Ecuación y= bx + a
Húmero H (mm)= 3.43·LT (mm) - 26.54
Radio H (mm)= 3.18·LT (mm) + 19.51 Sb
= 8.96
Ulna H (mm)= 2.78·LT (mm) + 6.21 Sb
= 7.12
Fémur H (mm)= 3.14·LT (mm) - 12.96
Tibia H (mm)= 2.92·LT (mm) + 9.41
Húmero + radio H (mm)= 1.65·LT [húmero+radio] (mm) - 4.32 Sb
= 4.03
Fémur + tibia H (mm)= 1.52·LT [fémur+tibia] (mm) - 2.47 Sb
= 3.95
I.3 K. M. Clark (1995) (n=15). Ecuación y= bx + a
Metacarpal II H (cm)= 0.94·LT (mm) - 1.56 r2
= 0.989
Metacarpal III H (cm)= 0.83·LT (mm) - 2.03 r2
= 0.993
Metacarpal IV H (cm)= 0.84·LT (mm) - 2.60 r2
= 0.994
Metacarpal V H (cm)= 0.98·LT (mm) - 1.56 r2
= 0.989
Metatarsal II H (cm)= 0.86·LT (mm) - 2.04 r2
= 0.995
Metatarsal III H (cm)= 0.77·LT (mm) - 2.26 r2
= 0.996
Metatarsal IV H (cm)= 0.75·LT (mm) - 2.68 r2
= 0.996
Metatarsal V H (cm)= 0.83·LT (mm) - 1.75 r2
= 0.996
I.4 Valadez Azúa (1998). Ecuaciones y= bx; y= x1
+ x2
+ x3
+ x4
Húmero H (mm)= 3.1·LT (mm)
Radio H (mm)= 3.3·LT (mm)
Tibia H (mm)= 2.9·LT (mm)
Extremidad anterior H (mm)= LT·[escápula + húmero + radio + metacarpal central (se usó el más largo)] (mm)
I.5 Valadez Azúa et al. (1998). Ecuación y= bx
Longitud cabeza- H (mm)= a·Lc-T 2
(mm); a= de 0.567ΘΘΘΘΘ1
a 0.628
tronco (ec.2)
I.6 Chrószcz et al. (2006) (n=91). Ecuación y= bx + a
Cavidad craneal H (cm)= 1.0067·LT (mm) - 32.227 Intervalo de confianza95%
= 2.56 r2
= 0.8216
II. Longitud (L)
II.1 G. R. Clark (1995, in Crockford 1997); longitud del cuerpo (LC
). Ecuación y= bx + a
Pelvis LC
(cm)= 0.47·LT (mm) - 15.7
Vértebras LC
(mm)ΘΘΘΘΘ2
= 1.04·L[13VT+7VL+VS (mm)] + 2.13
II.2 Valadez Azúa (1998); longitud cabeza-tronco, ec. 1 (Lc-T 1
). Ecuación y= x1
+ x2
Cráneo+vértebras Lc-T 1
(mm)= longitud máxima craneal (mm) + longitud atlas-sacro (mm)
II.3 Valadez Azúa et al. (1998); longitud cabeza-tronco, ec. 2 (Lc-T 2
). Ecuación y= x/b
Dentario Lc-T 2
(mm)= longitud de la rama horizontal (mm)/b; b= de 0.109ΘΘΘΘΘ3
a 0.119
III. Peso o masa (P)
III.1 Wing (1976, in Hamblin, 1984). Ecuación log y= b·log x + a
Dentario log P (g)= 2.1122·log (altura en el punto medio del M1 (mm)) + 1.2722
III.2 Wing (1978)
Dentario log P (g)= 2.2574·log (altura en el punto medio del M1 (mm)) + 1.1164 r = 0.9808
p1-p4 log P (g)= 3.0691·log (longitud hilera premolar inferior (mm)) - 0.7307 r = 0.948
m1-m3 log P (g)= 3.2735·log (longitud hilera molar inferior (mm)) - 0.8873 r = 0.934
Cuadro 1. Ecuaciones para estimar la alzada, la longitud del cuerpo y el peso en perros.](https://image.slidesharecdn.com/leer2-140316152627-phpapp01/85/arqueociencias-32-320.jpg)




![Arqueo·Ciencias 37
glosado el marco comparativo.
Dimorfismo sexual secundario. Finalmente,
se verificó la aplicabilidad de los criterios de di-
morfismo sexual secundario, tanto los morfológi-
cos (cuadro 3: The y Trouth, 1976; Shigehara et al,
1997), como los morfométricos (cuadro 2: Trouth
et al., 1977; Monterroso Rivas, 2004); se incluyeron
dos de los índices elaborados por Monterroso Ri-
vas (2004) y en los que los valores calculados
por ese autor para cada sexo no se solaparon,
aunque en esta ocasión, se invirtió la fórmula del
índice de las anchuras del paladar con la finalidad
de obtener las cifras que ese autor registra.
Resultados
Alzada (cuadro 4). Las ecuaciones de Valadez
Azúa son las que en promedio estiman la alzada
más baja (459.8 mm), siguiendo en orden ascen-
dente las de Koudelka (466.7 mm), la de Chrószcz
et al. (468 mm), las de Harcourt (471.7 mm), las
de K. M. Clark (481 mm), y la de Valadez Azúa y co-
laboradores (721.9 mm). Las alzadas tuvieron di-
ferencias de diversa magnitud entre las estima-
ciones máxima y mínima en cada serie, así como
diferente coeficiente de variación, obteniéndose
los valores más bajos con las ecuaciones de Har-
court (Koudelka, diferencia: 43.33 mm, CV=3.5;
Harcourt, 19.12 mm, CV=1.7; Valadez Azúa, 21.6
mm, CV=2.1; Clark, 3.4 mm, CV=2.4; Valadez Azúa
et al., 63.3 mm, CV=6.2).
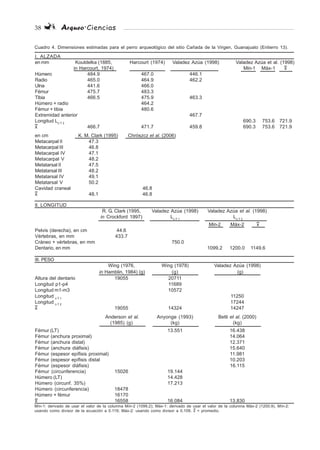
Carácter Macho Hembra
Ensanchamiento de la parte inferior Notoria No notoria
de la cresta frontal externa
Cresta sagital Bien desarrollada (elevada) a lo Poco desarrollada (baja), localizada
largo de los parietales y el sobre el interparietal
interparietal
Convergencia de las líneas Justo por detrás del bregma, Muy posterior al bregma
temporales formando la cresta sagital que se
continúa en el hueso interparietal
Tamaño y forma de la línea nucal No es recta y está curvada hacia Es recta
superior (región occipital) arriba
Cresta condiloidea de la mandíbula Muy marcada Tenue
Ángulo subpúbico de la pelvis Ángulo recto o ligeramente mayor Ángulo claramente obtuso
a 90°
Marca de inserción de los músculos Alargadas Redondeadas
rectus capitis ventralis major y
minor en el basioccipital*
Cuadro 3. Criteros propuestos para sexar restos de perro. Tomado de Shigehara et al. (1997) y The y Trouth (1977)*.
Sin contar la serie de Valadez y colaborado-
res, que se aparta del intervalo de 400 a 500 mm
en la que oscilan las otras estimaciones, los va-
lores difieren hasta 60 mm o 14 % [valor mínimo
(Kou-delka), 441.6 mm; valor máximo (Clark), 502
mm] y en conjunto generan un coeficiente de va-
riación de 2.7.
La prueba de Kruskal-Wallis indicó la exis-
ten-cia de diferencias significativas (H0.95,3
= 8.743,
P=0.03), debidas precisamente a los resultados
de las series de ValadezAzúa y de Clark (diferencia
de medias jerarquizadas, 12.5>Z= 9.6), y que son
justamente las series que generan los promedios
más bajo y más alto. Al omitir la estimación más
baja de Valadez Azúa o la más alta de Clark, las
estimaciones restantes pertenecen a la misma
población (valor inferior omitido, 446.1:
H0.95,3
=7.343, P=0.06; valor superior omitido: 502,
H0.95,3
=7.553, P=0.06; ambos valores omitidos:
H0.95,3
=5.979, P=0.11); es de notar que al eliminar-
se únicamente el valor mínimo de todo el conjunto
y que es generado por una de las ecuaciones de
Koudelka (441.6 mm), se incrementa la diferen-
cia de todo el conjunto, misma que es ocasiona-
da nuevamente por las series de Valadez Azua y
de Clark (H0.95,3
=9.22, P=0.03; diferencia de me-
dias jerarquizadas 12.5>Z=9.152). La alzada es-
timada con la única ecuación de Chrószcz et al.
(2006), 468 mm, queda inmersa en el intervalo
obtenido con los cuatro conjuntos de ecuaciones
analiza-das, por lo que podría considerarse que
pertenece a la misma población de resultados.](https://image.slidesharecdn.com/leer2-140316152627-phpapp01/85/arqueociencias-37-320.jpg)




![44 Arqueo·Ciencias
de las ciencias de la salud. Limusa Wiley, 4ª edición,
México. 75 pp. + apéndices e índice temático.
Del Pozo, E. C. 1984 [1985]. Obras completas 7: comen-
tarios a la obra de Francisco Hernández. Universidad
Nacional de México, México. 376 pp.
Desse, J. 1984. Propositions pour une réalisation colecti-
ve d’un corpus: fiches d’identificación et d’explotation
métrique du squelette des poisons. Pp. 67-86, in
2èmes rencontres d’archéoichthyologie (N. Desse-
Berset, ed.). Notes et monographies techniques, Édi-
tions du Centre National de la Recherche Scientifique,
París, 16:1-200.
Desse, J. y N. Desse-Berset. 1996. Archaeozoology of
groupers (Epinephelinae). Identification, osteometry
and keys to interpretation.Archaeofauna, 5:121-127.
Drake,A. G. y C. P. Klingenberg. 2010. Large-scale diver-
sification of skull shape in domestic dogs: disparity
and modulariy. TheAmerican Naturalist, 175(3):289-
301.
Evans, H. E. 1993. Miller’s anatomy of the dog. 3a
ed.,
Saunders. Philadelphia. 1113 pp.
Forest, V. 1998. De la hauteur au garrot des espèces
domestiques en archéozoologie. Revue du Médecine
Vétérinaire, 149:55-60.
García-Bárcena, J. 1982. Instructivo. Sección de Labora-
torios, Departamento de Prehistoria, Instituto Nacional
deAntropología e Historia, México. 59 pp.
Guzmán,A. F. y O. J. Polaco. 2000. Los peces arqueológi-
cosdelaOfrenda23delTemploMayordeTenochtitlan.
Colección Científica del Instituto Nacional deAntropo-
logía e Historia, 418:1-225.
Guzmán,A. F. y O. J. Polaco. 2002. Hiperostosis en pe-
ces mexicanos. Archaeofauna, 11:45-62.
Guzmán,A. F. y O. J. Polaco. 2003. El consumo de peces
en una casa del siglo XVI en la ciudad de México. Pp.
39-73, in Excavaciones del Programa deArqueología
Urbana (E. Matos Moctezuma, coord.). Colección
Científica del Instituto Nacional de Antropología e
Historia, México, 452:1-326.
Guzmán, A. F. y O. J. Polaco. 2005. La arqueoictiología
en México. Revista Digital Universitaria, 6(8): 10 pp.
http://www.revista.UNAM.mx/vol.6/num8/art85/
int85.htm.
Guzmán,A.F.yO.J.Polaco.2007.FishinginPre-hispanic
México. Pp. 25-36, in The role of fishing in ancient
time: Proceedings of the 13th
Meeting of the ICAZ-
Fish Remains Working Group Meeting (H. Hüster
Plogmann, hrsg.). Internationale Archäologie,
Arbeitsgemeinshcalt, Symposium, Tagung, Kongress,
Band 8. Verlag Marie Leidorf GmbH. Rahden, Westf.
176 pp.
Guzmán,A. F. y O. J. Polaco. 2008a.Análisis de los res-
tos de fauna del sitio Cañada de la Virgen, Guanajua-
to. Informe Z-589, Laboratorio de Arqueozoología,
Subdirección de Laboratorios y Apoyo Académico,
Instituto Nacional deAntropología e Historia, México.
12 pp.
Guzmán, A. F. y O. J. Polaco. 2008b. Faunal resources
from Chalco: early Toltec to late Aztec/Los recursos
faunísticos de Chalco: Tolteca temprano a Azteca
tardío. Pp. 303-326, in Un lugar de jade: sociedad y
economía en el antiguo Chalco/Place of jade: society
and economy in ancient Chalco (M. G. Hodge, ed.).
Instituto Nacional deAntropología e Historia y Univer-
sity of Pittsburgh, SerieArqueología de México/Latin
AmericanArchaeology Publications, México. 506 pp.
Guzmán,A. F. y Ó. J. Polaco. 2009. Peces fósiles mexica-
nos de agua dulce. Pp. 316-340, in Setenta y cinco
años de la Escuela Nacional de Ciencias Biológicas
(J. Ortega Reyes, J. E. Sedeño Díaz y E. López López,
comps.). Instituto Politécnico Nacional, México. 408
pp.
Guzmán,A. F., J.Arroyo-Cabrales y O. J. Polaco. 2006.
Colección Osteológica del Laboratorio deArqueozoo-
logía “M. en C. Ticul Álvarez Solórzano”, INAH. Pp.
347-357, in Colecciones mastozoológicas de México
(C. Lorenzo, E. Espinoza, M. Briones y F.A. Cervan-
tes, eds.). Instituto de Biología, UNAM, yAsociación
Mexicana de Mastozoología,A. C., México. 572 pp.
Guzmán,A.F.,O.J.PolacoyH.P.Pollard.2001.Ofrendas
de peces asociadas a entierros del Clásico-Epiclásico
en Urichu, Michoacán, México. Archaeofauna, 10:
149-162.
Hamblin, N. L. 1984.Animal use by the Cozumel Maya.
The University of Arizona Press, Tucson. 206 pp.
Handley, B. 2000. Preliminary results in determining dog
types from prehistoric sites in the Northeastern United
States. Pp. 205-215, inDogs through time: an archaeo-
logical perspective (S. J. Crockford, ed.). BAR Interna-
tional Series núm. 889,Victoria, Canadá. 343 pp.
Harcourt, R. A. 1974. The dog in prehistoric and early
historic Britain. Journal of Archaeological Science,
1:151-175.
Koudelka, F. 1885. Das Verhältnis der ossa longa zur
SkeletthöhebeidenSäugethieren.Verhandlungendes
Naturforschenden Vererenis in Brünn, 24:127-153.
Lawrence, B. 1967. Early domestic dogs. Sonderdruck
aus Z. F. Säugetierkunde, 32(1):44-59.
Lawrence, B. 1968. Antiquity of large dogs in North
America. Tewiba (The Journal of the Idaho State Uni-
versity Museum), 11(2):43-49.
Lawrence, B. y W. H. Bossert. 1967. Multiple characters
analysis of Canis lupus, latrans and familiaris with
a discussion of the relationships of Canis niger. Ame-
rican Zoologist, 7:223-232
Leach, F. yA. Boocock. 1995. Estimating live fish catches
from archaeological bone fragments of red snapper,
Pagrus auratus. Tuhinga: records of the Museum of
New Zealand Te Papa Tongarewa, 3:1-28.
León y Gama,A. de. 1990. Descripción histórica y crono-
lógica de las dos piedras. Pp. 41-95, in Trabajos ar-
queológicos en el centro de la Ciudad de México (E.
Matos Moctezuma, coord.).Antologías, SerieArqueo-
logía, Instituto Nacional deAntropología e Historia, 2ª
ed., México. 854 pp. Publicado originalmente en 1792.
Lignereux,Y., S. Regodon y C. I. Pavaux, 1991.Typologie
céphalique canine. Revue du Médicine Vétérinaire
142:469-480.](https://image.slidesharecdn.com/leer2-140316152627-phpapp01/85/arqueociencias-44-320.jpg)





























































