
Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Destacado (20)
Similar a Reproducción humana: formación de gametos masculinos
Similar a Reproducción humana: formación de gametos masculinos (20)
Más de albertososa
Más de albertososa (20)
Reproducción humana: formación de gametos masculinos
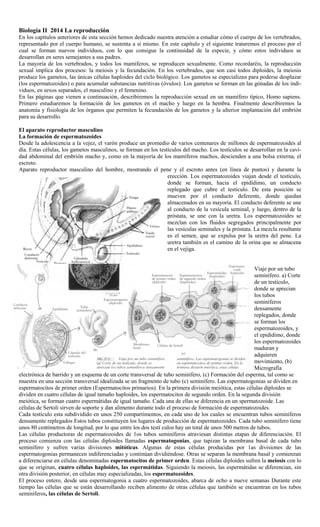
- 1. Biología II 2014 La reproducción En los capítulos anteriores de esta sección hemos dedicado nuestra atención a estudiar cómo el cuerpo de los vertebrados, representado por el cuerpo humano, se sustenta a sí mismo. En este capítulo y el siguiente trataremos el proceso por el cual se forman nuevos individuos, con lo que consigue la continuidad de la especie, y cómo estos individuos se desarrollan en seres semejantes a sus padres. La mayoría de los vertebrados, y todos los mamíferos, se reproducen sexualmente. Como recordaréis, la reproducción sexual implica dos procesos: la meiosis y la fecundación. En los vertebrados, que son casi todos diploides, la meiosis produce los gametos, las únicas células haploides del ciclo biológico. Los gametos se especializan para poderse desplazar (los espermatozoides) o para acumular substancias nutritivas (óvulos). Los gametos se forman en las gónadas de los indi- viduos, en sexos separados, el masculino y el femenino. En las páginas que vienen a continuación, describiremos la reproducción sexual en un mamífero típico, Homo sapiens. Primero estudiaremos la formación de los gametos en el macho y luego en la hembra. Finalmente describiremos la anatomía y fisiología de los órganos que permiten la fecundación de los gametos y la ulterior implantación del embrión para su desarrollo. El aparato reproductor masculino La formación de espermatozoides Desde la adolescencia a la vejez, el varón produce un promedio de varios centenares de millones de espermatozoides al día. Estas células, los gametos masculinos, se forman en los testículos del macho. Los testículos se desarrollan en la cavi- dad abdominal del embrión macho y, como en la mayoría de los mamíferos machos, descienden a una bolsa externa, el escroto. Aparato reproductor masculino del hombre, mostrando el pene y el escroto antes (en línea de puntos) y durante la erección. Los espermatozoides viajan desde el testículo, donde se forman, hacia el epidídimo, un conducto replegado que cubre el testículo. De esta posición se mueven por el conducto deferente, donde quedan almacenados en su mayoría. El conducto deferente se une al conducto de la vesícula seminal, y luego, dentro de la próstata, se une con la uretra. Los espermatozoides se mezclan con los fluidos segregados principalmente por las vesículas seminales y la próstata. La mezcla resultante es el semen, que se expulsa por la uretra del pene. La uretra también es el camino de la orina que se almacena en el vejiga. Viaje por un tubo seminífero. a) Corte de un testículo, donde se aprecian los tubos seminíferos densamente replegados, donde se forman los espermatozoides, y el epidídimo, donde los espermatozoides maduran y adquieren movimiento, (b) Micrografía electrónica de barrido y un esquema de un corte transversal de tubo seminífero, (c) Formación del esperma, tal como se muestra en una sección transversal idealizada se un fragmento de tubo (c) seminífero. Las espermatogonias se dividen en espermatocitos de primer orden (Espermatocitos primarios). En la primera división meiótica, estas células diploides se dividen en cuatro células de igual tamaño haploides, los espermatocitos de segundo orden. En la segunda división meiótica, se forman cuatro espermátidas de igual tamaño. Cada una de ellas se diferencia en un spermatozoide. Las células de Sertoli sirven de soporte y dan alimento durante todo el proceso de formación de espermatozoides. Cada testículo esta subdividido en unos 250 compartimentos, en cada uno de los cuales se encuentran tubos seminíferos densamente replegados Estos tubos constituyen los lugares de producción de espermatozoides. Cada tubo seminífero tiene unos 80 centímetros de longitud, por lo que entre los dos testí culos hay un total de unos 500 metros de tubos. Las células productoras de espermatozoides de 1os tubos seminíferos atraviesan distintas etapas de diferenciación. El proceso comienza con las células diploides llamadas espermatogonias, que tapizan la membrana basal de cada tubo seminífero y sufren varias divisiones mitóticas. Algunas de estas células producidas por 1as divisiones de las espermatogonias permanecen indiferenciadas y continúan dividiéndose. Otras se separan la membrana basal y comienzan a diferenciarse en células denominadas espermatocitos de primer orden. Estas células diploides sufren la meiosis con lo que se originan, cuatro células haploides, las espermátidas. Siguiendo la meiosis, las espermátidas se diferencian, sin otra división posterior, en células muy especializadas, los espermatozoides. El proceso entero, desde una espermatogonia a cuatro espermatozoides, abarca de ocho a nueve semanas Durante este tiempo las células que se están desarrollando reciben alimento de otras células que también se encuentran en los tubos seminíferos, las células de Sertoli.
- 2. El trayecto del espermatozoide El camino que sigue el espermatozoide en su viaje hacia fuera se puede seguir en la figura. Desde los testículos, los espermatozoides son conducidos hacia el epidídimo, que consta de un tubo largo y replegado sobre el testículo. Se halla rodeado por una capa fina de fibras musculares lisas. Los espermatozoides aún no son móviles en el momento de alcanzar el epidídimo, y sólo poseen motilidad unos 12 días, en promedio, después de haberse concentrado allí a) Imagen frontal el aparato masculino, mostrando el camino que siguen los espermatozoides en su recorrido por los conductos, (b) Sección transversal del pene humano. El pene está formado por tres masas cilindricas de tejido eréctil esponjoso que contiene un gran número de pequeños espacios, cada uno del tamaño de una cabeza de aguja. La erección del pene está provocada por la dilatación de las arterias que transportan la sangre hacia los tejidos esponjosos y la compresión de las venas que salen de estos tejidos. Desde el epidídimo, el espermatozoide va hacia el conducto deferente, donde se almacena la mayoría de ellos. Cada conducto deferente, prolongación de los tubos replegados del epidídimo, conectan cada testículo a la cavidad abdominal. Los conductos deferentes están recubiertos de tres capas de musculatura lisa, cuyas contracciones empujan a los espermatozoides a lo largo conducto. En la pared posterior de la cavidad abdominal, los conductos deferentes rodean la vejiga y después se unen con los conductos de las vesículas seminales. El conducto deferente de cada testículo entra en la próstata y se une con la uretra, que se prolonga internamente por el pene. La uretra sirve como conducto para expulsar la orina y para la descarga rápida del semen durante la eyaculación, procesos ambos que nunca pueden producirse a la vez. Mientras los espermatozoides van siguiendo su camino por los conductos, se van sumando cantidades variables de fluidos procedentes de las vesículas seminales y de la próstata. Las vesículas seminales segregan un líquido con abundante fructosa que nutre a los espermatozoides. El líquido contiene también una elevada concentración de prostaglandinas, las cuales estimulan las contracciones en la musculatura del aparato reproductor femenino. Estas contracciones ayudan a los espermatozoides a continuar su viaje hacia el óvulo. La próstata aporta un fluido alcalino lechoso que ayuda a neutralizar el ambiente ácido del conducto reproductor femenino. La suspensión de espermatozoides en el fluido que hemos comentado constituye el semen. En cada eyaculación se liberan entre 3 a 4 mililitros de semen. Erección del pene y orgasmo masculino La función del pene es depositar los espermatozoides dentro del aparato reproductor femenino. La erección del pene, que puede producirse por una serie de estímulos, se produce por el aumento del flujo sanguíneo en sus tejidos esponjosos eréctiles. El flujo de sangre se controla por las terminaciones nerviosas que inervan las arterias del tejido eréctil. Cuando los tejidos se dilatan, las venas se comprimen y por lo tanto se evita que la sangre marche del tejido. Con una estimulación continuada, el pene se engrosa y se endurece. La erección va acompañada por la descarga en la uretra de una pequeña cantidad de líquido procedente de las glándulas bulbouretrales, dos pequeñas estructuras situadas en la base del pene. Este fluido sirve de lubrificante, ayudando al movimiento del esperma por la uretra y a la penetración del pene en la hembra. La estimulación continuada de los receptores sensitivos del pene envían una serie de impulsos nerviosos ascendentes a través de arcos reflejos de la parte inferior de la médula espinal a las neuronas motoras que inervan los diferentes músculos del aparato reproductor. La contracción de la musculatura lisa de las paredes de los conductos deferentes causan que el esperma se vacíe en la uretra. Otros músculos ayudan empujar el semen por la uretra y expulsarlo del pene. Estos espasmos musculares que se producen durante la eyaculación, dan origen a gran parte de las sensaciones asociadas al orgasmo. El papel de las hormonas Además de los espermatozoides, los testículos son también la principal fuente de hormonas masculinas, que colectivamente se denominan andrógenos. El andrógeno más importante es la testosterona, una hormona esteroidea necesaria para la maduración de los espermatozoides. Está producida especialmente por las células intersticiales, que se encuentran en el tejido que rodea los tubos seminíferos de los tejidos. La corteza de la cápsula suprarrenal segrega también pequeñas cantidades de andrógenos. Los andrógenos comienzan ya a fabricarse durante el desarrollo embrionario, haciendo que el feto se desarrolle como macho y no como hembra. Después del nacimiento, la producción de andrógenos continúa produciéndose en muy pequeñas cantidades hasta que el niño alcanza los 10 años de edad aproximadamente. Desde entonces, se inicia un aumento en su secreción y comienza la producción de espermatozoides. Se marca así el inicio de la pubertad, que va acompañada de un aumento de los tejidos y también de la próstata y otros órganos accesorios. La testosterona también afecta las partes del cuerpo que no están directamente implicadas en la producción y deposición de los espermatozoides. En el macho de la especie humana, estos efectos incluyen el crecimiento de la laringe y el cambio de la voz más profunda, el crecimiento del esqueleto y la aparición de pelo en partes características. Los andrógenos estimulan la síntesis de proteínas y, por lo tanto, del tejido muscular. También estimulan las glándulas sudoríparas apocrinas, cuyas secreciones favorecen a las bacterias y producen los olores del cuerpo asociados con el sudor después de la pubertad. También pueden producir una hiperactividad de las glándulas sebáceas, dando origen al acné. Estas características asociadas con las hormonas, aunque no directamente con la reproducción, se conocen como caracteres
- 3. secundarios En otros animales, la testosterona es responsable de los caracteres sexuales secundarios como las astas del reno, la melena del león, la poderosa musculatura y el porte fiero del caballo garañón, la cresta y espolones del gallo y el plumaje vistoso de muchos machos de aves. Es también responsable de las pautas de comportamiento como la señalización olorosa que realizan los perros o todas las formas de agresión que se dirigen hacia otros machos en una gran variedad de especies de vertebrados. La producción de hormonas que controlan el funcionamiento del testículo está regulada por un sistema de retroacción negativa. La hormona producida en el hipotálamo, GnRH o factor de liberación gonadotrópico, estimula la hipófisis anterior para que produzca la hormona luteinizante (LH) y la hormona estimulante de los folículos (FSH). La LH estimula la producción y la liberación de testosterona por las células intersticiales del testículo. A continuación, la producción de GnRH por el hipotálamo queda inhibida por los niveles de testosterona. Se cree que la testosterona también influye sobre la hipófisis suprimiendo la producción de LH. Como consecuencia de estos efectos inhibidores combinados, la secreción de LH por la hipófisis se reduce. La FSH actúa sobre las células de Sertoli, las cuales producen, a su vez, la hormona inhibina. Esta hormona, realizando otra vía de retroactividad negativa, inhibe la formación de FSH. El efecto combinado de la testosterona y de FSH es esencial para la producción de los espermatozoides. Regulación de la producción hormonal La producción de testosterona está regulada por un sistema de retroacción negativa en la que participa la hormona gonadotrópica llamada hormona luteinizante (LH). La hormona luteinizante es segregada en la hipófisis anterior, bajo la influencia del factor de liberación hormonal de la gonadotropina (GnRH), hormona segregada en el hipotálamo. la LH es transportada por la sangre hacia las células intersticiales de los tejidos, donde allí estimula la producción de testosterona. A medida que la concentración de testosterona en sangre aumenta la producción de LH por la hipófisis disminuye. Los testículos están también bajo la influencia de una hormona hipofisiaria, la hormona estimulante de los folículos (FSH). Actúa sobre las células de Sertoli de los testículos, y a través de ellas, sobre el desarrollo de los espermatozoides. Entre los factores que influyen en la regulación de la producción de FSH se encuentra una hormona de tipo proteico, la inhibina, segregada por las células de Sertoli. Esta hormona inhibe la producción de FSH. En el hombre, la secreción de testosterona es bastante constante. Sin embargo, en muchos machos de otros animales la producción hormonal cambia estacionalmente, influida por los cambios de temperatura, las horas de luz u otros cambios ambientales. La producción de la testosterona puede también quedar influida por circunstancias sociales. Los estudios realizados en toros, por ejemplo, han demostrado que después de ver una vaca, su concentración de LH llega a incrementarse hasta siete veces, y en media hora la concentración llega a un máximo. En muchas sociedades de animales, como los lobos o los licaones (perros cazadores de África), los machos socialmente inferiores nunca alcanzan la madurez sexual, presumiblemente porque la producción de testosterona se encuentra deprimida. La producción de testosterona humana puede variar igualmente por el ambiente emocional. En un estudio realizado entre las fuerzas armadas de Estados Unidos durante la guerra del Vietnam, los niveles de testosterona en los reclutas que se entrenaban y en las tropas de combate eran marcadamente bajos en comparación con los hombres situados en posiciones alejadas del frente. La producción de testosterona está también muy influida por los compuestos sintéticos conocidos como esteroides anabolizantes. Estos productos, variaciones químicas de la hormona testosterona, comenzaron a usarse en un principio en Alemania, durante la década de 1930, en el intento de producir el crecimiento muscular sin los efectos de la masculinización. Debido a su semejanza con la hormona testosterona, los anabolizantes funcionan como inhibidores en el sistema de retroacción negativa que regula la producción de testosterona. En los varones, su utilización hace caer la producción de testosterona en un 85 por ciento, provocando la reducción de testículos y crecimiento de pechos. El uso prolongado de estos compuestos puede hacer aumentar enormemente el riesgo de lesión renal o hepática, cáncer de hígado o fallo cardíaco. En los adolescentes, los esteroides anabolizantes pueden provocar calvicie prematura y detención del crecimiento. El aparato reproductor femenino El aparato reproductor femenino se representa en la Figura 37-8. Los ovarios, de unos 3 centímetros de diámetro, son los órganos productores de gametos. Están formados por masas compactas de células, suspendidas en la cavidad abdominal por una banda de tejido conjuntivo. Los ovocitos, de los que se desarrollarán los óvulos, se encuentran en la capa más externa del ovario. Otras estructuras importantes del aparato reproductor femenino son las trompas (oviductos), el útero, la vagina y la vulva. El útero es un órgano muscular hueco, piriforme de unos 7,5 centímetros de largo por 5 centímetros de ancho en las mujeres no embarazadas. Su posición es casi horizontal, en la cavidad abdominal, por encima de la vejiga urinaria. La
- 4. capa interna del útero, llamada endome-trio, tiene dos capas principales, una de las cuales se expulsa durante la menstruación y la restante, la capa más profunda, sirve para regernerar la que se ha expulsado. La musculatura lisa de las paredes del útero se mueven siguiendo ondas continuas. Este movimiento favorece el movimiento de los espermatozoides hacia las trompas, y el del ovocito que se desarrolla en óvulo cuando se dirige desde las trompas hacia el útero. Estas contracciones aumentan cuando el endometrio es expulsado durante el período menstrual, y se producen al máximo cuando la mujer da a luz. El músculo circular que se encuentra en la apertura del útero se llama cérvix. Los espermatozoides pasan por esta abertura en su camino hacia los ovocitos. Cuando llega el momento del parto, el cérvix se dilata para permitir la expulsión del feto. La vagina es un tubo muscular de unos 8 a 10 centímetros de largo que conecta la cérvix con el exterior del cuerpo. Es un órgano receptivo para el pene y también constituye el canal de nacimiento. El orificio exterior se sitúa entre el orificio urinario y el ano. Los órganos sexuales externos se denominan en conjunto vulva. El clítoris, la mayor parte del cual queda rodeado por la piel circundante, es el órgano de unos dos centímetros, homólogo al pene del macho (en una etapa inicial embrionaria, no existe diferencia entre sexos). Los labios son pliegues de la piel. Los labios mayores son gruesos y, en edad adulta, están cubiertos de vello púbico. Cubren y protegen las estructuras internas más delicadas, entre las que destacan los labios menores, más membranosos. Formación de los ovocitos En las hembras, los ovocitos de primer orden empiezan a formarse en el tercer mes de vida embrionaria. En el momento que nace una niña, sus dos ovarios llegan a contener unos 2 millones de ovocitos primarios, que son todos los que poseerá en su vida. Estas células diploides se encuentran en un estado de profase de la primera división meiótica, y permanecen en este estadio hasta que la mujer no madura sexualmente. Entonces, bajo la influencia de las hormonas, la primera división meiótica del ovocito primario se realiza completamente. De los 2 millones de ovocitos que originalmente poseía la mujer, entre 300 y 400 consiguen alcanzar la madurez, uno cada 28 días, desde la pubertad hasta la menopausia, que típicamente se produce de camino a los 50 años de edad. Dando estas cifras, se puede afirmar que algunos ovocitos tardan más de 50 años desde el comienzo de la primera división meiótica hasta el fin de la misma. Cuando un ovocito primario está a punto de acabar la meiosis, su envoltura nuclear se fragmenta y los cromosomas se dirigen hacia la superficie celular. Al dividirse el núcleo, el citoplasma del ovocito se abulta. Un juego de cromosomas se dirigen hacia el abultamiento, que entonces se estrangula y se separa en forma de una pequeña célula llamada corpúsculo polar primario. El resto del material celular forma el ovocito de segundo orden (ver Figura 11-11, página 198). La primera división meiótica se finaliza en pocas horas antes de producirse la ovulación (expulsión del ovocito del ovario). La segunda división meiótica no se produce hasta que se alcanza la fecundación. Esta división produce el óvulo y otro corpúsculo basal diminuto. Los cuerpos polares se desintegran y sus núcleos, de hecho, se destruyen. Para que el ovocito madure es necesario que se produzca la meiosis y el aumento considerable de su tamaño. El aumento en diámetro refleja, de hecho, que se produce una acumulación de materiales de reserva y otras utilidades metabólicas, como el RNA mensajero y las enzimas, necesarias para las primeras etapas del desarrollo embrionario. Como consecuencia de las divisiones desiguales de la meiosis, la mayor parte de los materiales de reserva del ovocito se transfieren a un solo óvulo. Esta célula única es muy grande, de unos 100 micrómetros de diámetro en la especie humana. Los ovocitos se desarrollan cerca de la superficie del ovario. Un ovocito en crecimiento y las células especializadas que lo rodean constituyen el folículo ovárico (Figura 37-9). Las células del folículo aportan alimento al ovocito en desarrollo y segregan también estrógenos, las hormonas que estimulan el crecimiento continuado del folículo e inician la regeneración del endometrio. Durante la etapa final de crecimiento, el folículo se desplaza hacia la pared del ovario y produce una pequeña protuberancia como si se tratara de una ampolla que finalmente revienta, expulsando el ovocito (Figura 37-10). Normalmente empiezan a desarrollarse varios folículos de una vez, pero sólo uno madura lo suficiente para liberar el ovocito, y los restantes degeneran. El trayecto del ovocito Cuando el ovocito se expulsa del folículo durante la ovu- lación, es arrastrado hacia la trompa próxima (el oviducto) mediante el movimiento de la trompa sobre la superficie del
- 5. ovario y por el impulso que transmiten los cilios que se sitúan sobre las proyecciones digitiformes de ésta sobre el ovario (Figura 37-11). Este mecanismo es tan eficaz que cuando una mujer tiene un solo ovario y una sola trompa, y ambos se encuentran en posiciones opuestas, puede quedarse embarazada. El ovocito se dirige entonces lentamente por la trompa, impulsado por ondas peristálticas producidas por la musculatura lisa de las paredes de la trompa. El viaje desde el ovario hasta el útero dura unos tres días. Aunque un ovocito no fecundado vive sólo unas 72 horas después de ser lanzado del folículo, parece ser que sólo puede ser fecundado dentro de la mitad de este tiempo. Por lo tanto, de ocurrir, la fecundación se produce en la trompa. Si el óvulo se fecunda, el embrión joven se implanta en el endometrio tres o cuatro días después de que alcance el útero, unos seis o siete días después de la fecundación. Si el ovocito no se fecunda, muere y la pared del endometrio del útero se desprende durante la menstruación. La esterilización de la mujer se consigue normalmente con una operación en las trompas, impediendo que los espematozoides alcancen el ovocito. La operación se llama ligadura de trompas. El movimiento de los ovocitos y de los espermatozoides también puede quedar alterado por causas naturales, por ejemplo, con una cicatriz después de haberse producido una infección. Hasta hace poco, era virtualmente imposible que mujeres con estas obstrucciones pudieran quedar embarazadas. Mediante el proceso de fecundación in vitro, sin embargo, el ovocito se extrae quirúrgicamente del ovario en el momento antes de la ovulación, se fecunda con el semen del marido en una cápsula del laboratorio y luego se inserta en el útero en el momento que hubiera tenido que llegar en caso de una fecundación normal en las trompas. Entre un 15 y un 20 por ciento de los casos en que se ha intentado la fecundación in vitro, el óvulo sobrevive a la manipulación y se desarrolla normalmente en un embrión. Orgasmo femenino Bajo la influencia de diversos estímulos, el clítoris y tejidos asociados se congestionan y se distienden por el aporte de sangre, tal como ocurre con el pene del varón. Este proceso, sin embargo, es algo más lento que en el hombre. La distensión de los tejidos va acompañado de una secreción en la vagina que lubrifica sus paredes y neutraliza su ambiente normalmente ácido. Las condiciones acidas en la vagina, no sólo son inhóspitas para las bacterias (para evitar posibles infecciones), sino también para los espermatozoides. El orgasmo en la mujer, como en el varón, está con- dicionado por contracciones musculares rítmicas, seguidas de la expulsión a las venas de la sangre de los tejidos congestionados. Músculos homólogos producen el orgasmo en los dos sexos, aunque en la mujer no hay eyaculación de fluido por la uretra ni por la vagina. En el orgasmo, el cérvix o cuello del útero desciende por la parte superior de la vagina, donde el semen tiende a concentrarse. El orgasmo de la hembra puede producir también la contracción de la trompas, lo que ayuda a empujar los espermatozoides hacia arriba. Sin embargo, el orgasmo femenino no es necesario para que se produzca la concepción. Regulación hormonal en la hembra: El ciclo menstrual La producción de ovocitos en todas las hembras de verte- brados es un proceso cíclico. Intervienen una interrelación de hormonas, cambios en las células foliculares y, en los mamíferos marsupiales y placentarios, cambios en el reves- timiento interno del útero. En la mujer, esta variación de las concentraciones de hormonas y cambios que se producen en los tejidos se conoce por el ciclo menstrual. El ciclo menstrual se halla programado y regulado por el hipotálamo. Las hormonas participantes en este com- plicadísimo sistema de retroacción que regula el ciclo son los estrógenos y progesterona (las hormonas sexuales femeninas), las gonadotropinas hipofisiarias LH y FSH y la hormona liberadora de gonadotropinas (GnRH) del hipotálamo. Esquema de todos los acontecimientos que se producen en un ciclo menstrual, implicando cambios de concentración hormonal, y anatómicos en el ovario y en la pared interna del útero (endometrio). El ciclo comienza con el primer día con flujo menstrual, causado por el desgarro del endometrio, el revestimiento interno del útero. El aumento de la concentración de FSH y LH al comenzar el ciclo estimula a que un folículo ovárico crezca y secrete estrógenos. Bajo la influencia de los estrógenos, el endometrio se regenera. El brusco aumento de la concentración de estrógenos antes de alcanzar la mitad del ciclo dispara una subida súbita de LH desde la hipófisis, lo que produce la expulsión del ovocito (ovulación). (Se desconoce el papel, si es que lo tiene, del aumento instantáneo de FSH.) Después de la ovulación, la concentración tanto de LH como FSH cae. El folículo ahora se convierte en el cuerpo lúteo, que produce progesterona y estrógenos. La progesterona continúa estimulando el endometrio, preparándose para la implantación del óvulo fecundado. Si la fecundación no se produce, el cuerpo lúteo degenera, la producción de progesterona entonces se detiene y el endometrio comienza a destruirse, las concentraciones de LH y FSH vuelven a subir y un nuevo ciclo comienza de nuevo Al comienzo del ciclo, que tomamos como primer día del flujo menstrual, los niveles de hormonas son bajos (Figura 37- 12). Después de unos pocos días, un ovocito y su folículo comienzan a madurar bajo la influencia de la FSH y la LH. A medida que se agranda el folículo, segrega cantidades superiores de estrógenos, que estimulan la regeneración del endometrio en preparación de la implantación de un óvulo fecundado. El rápido aumento de las concentraciones de estrógeno cerca de la mitad del ciclo hace disparar también la secreción de LH por la hipófisis. Este pico de LH estimula a que el folículo expulse el ovocito e inicie su andadura hacia el útero. Bajo el estímulo continuado de la LH, las células del folículo ya vacío crecen y rellenan la cavidad dejada, formándose el cuerpo lúteo (o amarillo), como se muestra en la Figura 37-9b. Cuando las células del cuerpo lúteo aumentan de tamaño,
- 6. empiezan a sintetizar cantidades importantes de progesterona, así como continúa la de estrógenos. Al aumentar los niveles hormonales de estrógenos y progesterona, se inhibe por su efecto la producción de GnRH del hipotálamo, y consiguientemente se inhibe la producción de LH y FSH por la hipófisis. Si la fecundación no se ha producido, el cuerpo lúteo se reabsorbe con lo que la producción de hormonas cae en picado, y el suministro de sangre hacia el endometrio se ve muy reducido. Sin el sustento hormonal y nutricio, el endometrio ya no puede automantenerse y una porción de él es expulsado junto con el fluido menstrual. Es entonces, que, en respuesta a la escasa concentración de hormonas ovári-cas, la producción de FSH y LH de la hipófisis comienzan a aumentar, seguida del siguiente desarrollo de un nuevo folículo y de la subida de estrógenos que marca el inicio de un nuevo ciclo. El ciclo menstrual dura unos 28 días, aunque existe bastante variación individual. Incluso en mujeres cuyos ciclos coinciden con la media, la ovulación no siempre se produce en el mismo momento del ciclo (esta es una razón por la que el «método del ritmo» no es un buen sistema de control de natalidad). El inicio del primer ciclo menstrual marca el comienzo de la pubertad en la niña. En promedio, el comienzo de la pubertad es de los 12,3 años, aunque en realidad pueden haber bastantes años de variación. La mayor producción de hormonas sexuales femeninas antes de la pubertad induce al desarrollo de los caracteres sexuales secundarios como la forma de las caderas y el crecimiento de los senos. Métodos anticonceptivos Sin usar ninguna técnica anticonceptiva, el 80 por ciento de las mujeres en edad fértil y con relaciones sexuales normales quedan embarazadas al año. Sin embargo, existen una gran variedad de métodos anticonceptivos para las parejas que deciden evitar o retrasar el embarazo. En la tabla 37-1, se resumen por orden de efectividad, en términos de número de embarazos medio por año, en mujeres de edad fértil que los utilizan. En la mayoría de casos se dan dos cifras en la columna de efectividad. La primera, más pequeña, es un número ideal, obtenido cuando el método se usa adecuadamente. La segunda cifra es un valor promediado, reflejando la experiencia real. Durante muchos años los métodos más usados habían sido los de barrera, como diafragmas y preservativos. En los años 60 y 70, muchas parejas abandonaron estos métodos mecánicos y se decantaron por la pildora, una combinación sintética de estrógenos y progesterona. Cuando se ingieren diariamente, hacen constante la concentración de estas hormonas por lo que detienen la producción de FSH y LH por la hipófisis. Sin FSH, los folículos ováricos no maduran, y en ausencia de LH, no se produce la ovulación, por lo que el embarazo es imposible. Sin embargo, recientemente se están imponiendo de nuevo los diafragmas y preservativos. En combinación con las cremas espermicidas, estos dos métodos, y especialmente el preservativo de látex, proporcionan una barrera no sólo para el esperma, sino también para numerosos microorganismos patógenos. Como revela la Tabla , su protección, en cambio no es absoluta.