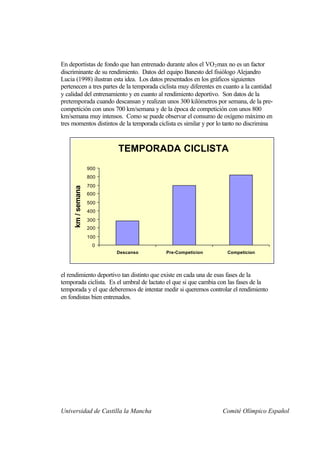
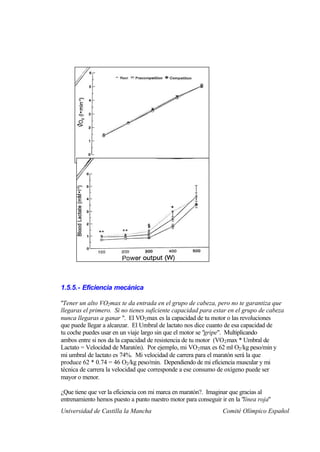
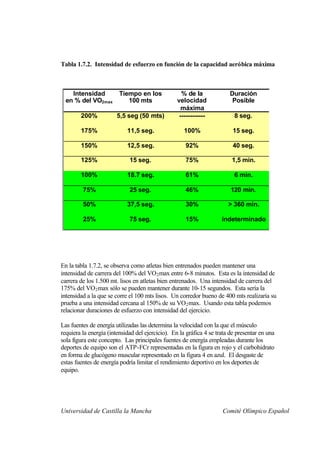
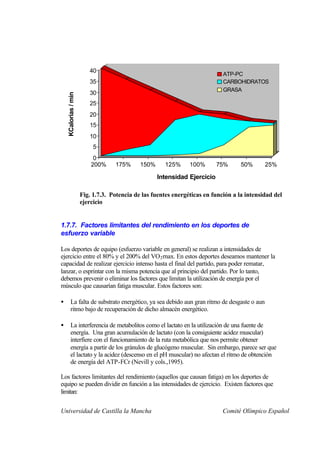
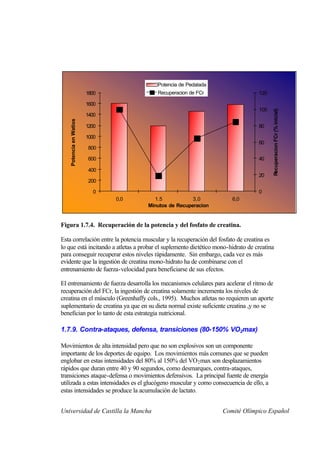
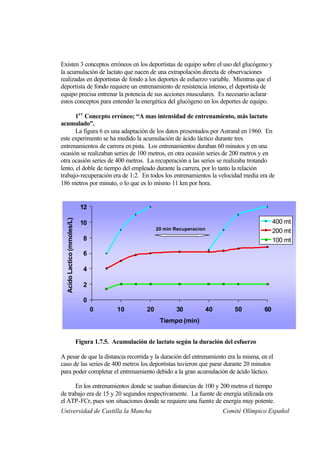
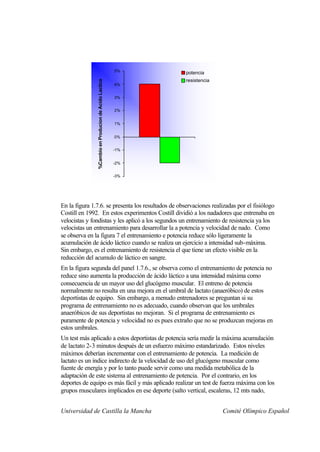
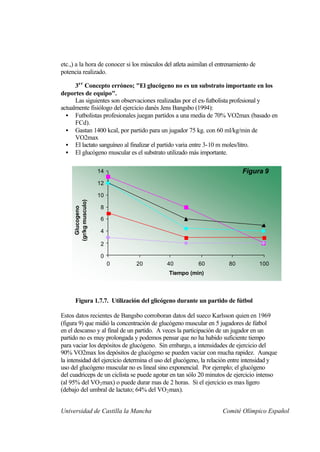
Este documento describe la fisiología del ejercicio a nivel celular y del sistema neuromuscular. Explica la estructura y función de las células, los tipos de células en el cuerpo como las musculares y nerviosas, y los componentes celulares como el ADN, membrana y organelos. También describe la estructura de las neuronas motoras, la unidad motora y los mecanismos de contracción muscular a nivel de proteínas y sarcómero.
























![Por lo tanto solamente 70% del volumen respiratorio ventila los alvéolos. Por lo tanto
seria mas lógico usar en vez de ventilación pulmonar (volumen respiratorio * frecuencia
respiratoria), ventilación alveolar ([volumen respiratorio – espacio muerto] * frecuencia
respiratoria) como un índice funcional de ventilación.
A la misma ventilación pulmonar, la ventilación alveolar será mayor cuanto mas
profundas sean las respiraciones (mayor volumen respiratorio). Contrariamente a lo que
veíamos con la eficiencia del sistema cardio-vascular y el aumento de la frecuencia
cardiaca, el aumento de la frecuencia respiratoria resulta en una ventilación alveolar
menos eficiente.
Universidad de Castilla la Mancha Comité Olímpico Español](https://image.slidesharecdn.com/fisiologadelejercicio-130408121532-phpapp01/85/Fisiologia-del-ejercicio-30-320.jpg)