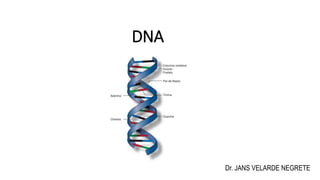
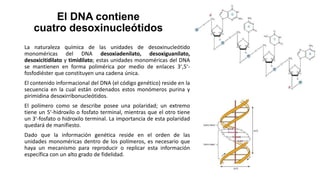
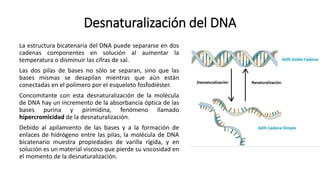
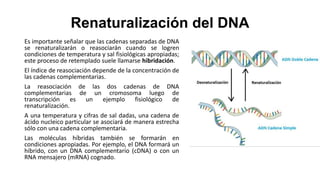
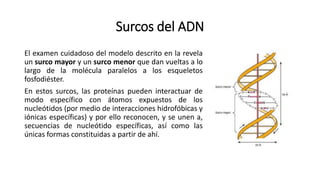
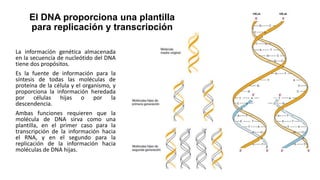
El documento describe la estructura y función del ADN. Explica que el ADN contiene la información genética codificada en la secuencia de cuatro nucleótidos y forma una doble hélice. También describe que el ADN sirve como plantilla para la replicación celular y la transcripción de genes en ARN.