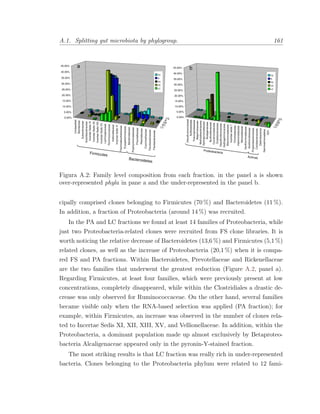
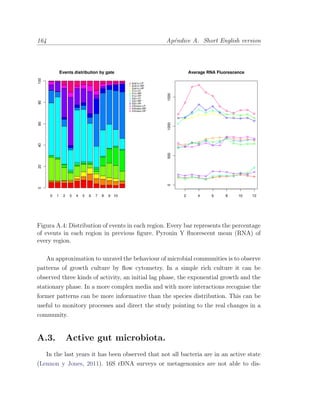
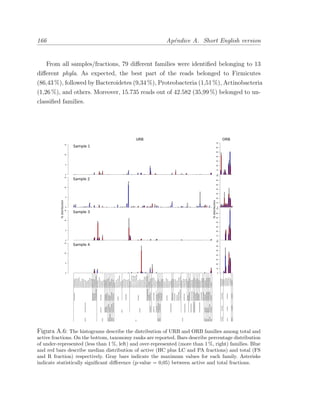
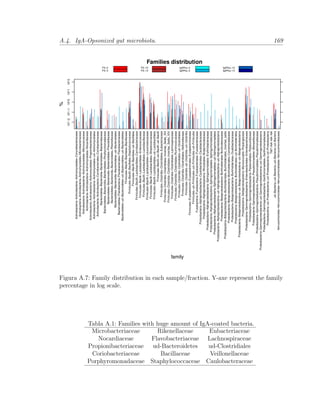
Este documento presenta la memoria de tesis doctoral de Francesc Peris-Bondia sobre la caracterización de la microbiota gastrointestinal humana mediante citometría de flujo y secuenciación de ADN. El objetivo principal es fraccionar poblaciones bacterianas complejas en función de marcadores celulares y estudiar su composición. Adicionalmente, se pretende identificar la fracción activa de la microbiota y estudiar su interacción con el sistema inmune a través de bacterias opsonizadas con inmunoglobulina A. La tesis aportará una visión más























































































































![Bibliograf´ıa 127
CAMPIERI, M. Bacteria as the cause of ulcerative colitis. Gut, 48(1):132–135, 2001. ISSN 00175749.
doi:10.1136/gut.48.1.132. URL http://gut.bmj.com/cgi/doi/10.1136/gut.48.1.132.
Cani, P. D; Neyrinck, A. M; Fava, F; Knauf, C; Burcelin, R. G; et al. Selective increases of
bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a me-
chanism associated with endotoxaemia. Diabetologia, 50(11):2374–83, 2007. ISSN 0012-186X.
doi:10.1007/s00125-007-0791-0. URL http://www.ncbi.nlm.nih.gov/pubmed/17823788.
Cani, P. D y Delzenne, N. M. The Role of the Gut Microbiota in Energy Metabolism and
Metabolic Disease. Current Pharmaceutical Design, 15(13):1546–1558, 2009. ISSN 13816128.
doi:10.2174/138161209788168164. URL http://openurl.ingenta.com/content/xref?genre=
article&issn=1381-6128&volume=15&issue=13&spage=1546.
Carlstedt-Duke, B y Lingaas, E. Influence of antibiotics on intestinal mucin in healthy subjects.
European Journal of, 1986. URL http://www.springerlink.com/index/U56172X1252G7502.
pdf.
Cazzolla, A. P; Campisi, G; Lacaita, G. M; Cuccia, M. A; Ripa, A; et al. Changes in pharyn-
geal aerobic microflora in oral breathers after palatal rapid expansion. BMC oral health, 6:2,
2006. ISSN 1472-6831. doi:10.1186/1472-6831-6-2. URL http://www.pubmedcentral.nih.
gov/articlerender.fcgi?artid=1388214&tool=pmcentrez&rendertype=abstract.
Cebra, J. Influences of microbiota on intestinal immune system development. The American journal
of clinical nutrition, 1999. URL http://www.ncbi.nlm.nih.gov/pubmed/10232647.
Chambers, S y Kunin, C. The osmoprotective properties of urine for bacteria: the protective effect
of betaine and human urine against low pH and high concentrations of electrolytes, sugars, and
urea. Journal of Infectious Diseases, 152(6):1308, 1985. URL http://jid.oxfordjournals.
org/content/152/6/1308.short.
Chaudhuri, R; McMahon, A. D; McSharry, C. P; Macleod, K. J; Fraser, I; et al. Serum and sputum
neurotrophin levels in chronic persistent cough. Clinical and experimental allergy : journal of the
British Society for Allergy and Clinical Immunology, 35(7):949–53, 2005. ISSN 0954-7894. doi:
10.1111/j.1365-2222.2005.02286.x. URL http://www.ncbi.nlm.nih.gov/pubmed/16008683.
Chiller, K; Selkin, B. a; y Murakawa, G. J. Skin microflora and bacterial infections of the skin.
The journal of investigative dermatology. Symposium proceedings / the Society for Investiga-
tive Dermatology, Inc. [and] European Society for Dermatological Research, 6(3):170–4, 2001.
ISSN 1087-0024. doi:10.1046/j.0022-202x.2001.00043.x. URL http://www.ncbi.nlm.nih.gov/
pubmed/11924823.](https://image.slidesharecdn.com/0ed7e562-4fd2-456a-857e-89fd1b64755c-161012124743/85/tesis-143-320.jpg)












![140 Bibliograf´ıa
ISSN 1340-2838. doi:10.1093/dnares/dsm018. URL http://www.pubmedcentral.nih.gov/
articlerender.fcgi?artid=2533590&tool=pmcentrez&rendertype=abstract.
La Duc, M. T; Dekas, A; Osman, S; Moissl, C; Newcombe, D; et al. Isolation and charac-
terization of bacteria capable of tolerating the extreme conditions of clean room environ-
ments. Applied and environmental microbiology, 73(8):2600–11, 2007. ISSN 0099-2240. doi:
10.1128/AEM.03007-06. URL http://www.pubmedcentral.nih.gov/articlerender.fcgi?
artid=1855582&tool=pmcentrez&rendertype=abstract.
Lander, E. S; Linton, L. M; Birren, B; Nusbaum, C; Zody, M. C; et al. Initial sequencing and analysis
of the human genome. Nature, 409(6822):860–921, 2001. ISSN 0028-0836. doi:10.1038/35057062.
URL http://www.ncbi.nlm.nih.gov/pubmed/11237011.
Lane, D. D. J; Stackenbrandt, E; y Goodfellow, M. 16S/23S rRNA sequencing. John Wiley and
Sons, nucleic ac ed´on
., 1991.
Larson, H; Price, A; y Honour, P. Clostridium difficile and the aetiology of pseudomembra-
nous colitis. The Lancet, 1978. URL http://linkinghub.elsevier.com/retrieve/pii/
s0140-6736(78)90912-1.
Lawton, J. H; Daily, G; y Newton, I. Population Dynamic Principles [and Discussion]. Philosophical
Transactions of the Royal Society B: Biological Sciences, 344(1307):61–68, 1994. ISSN 0962-8436.
doi:10.1098/rstb.1994.0052. URL http://rstb.royalsocietypublishing.org/cgi/doi/10.
1098/rstb.1994.0052.
Lay, C; Rigottier-Gois, L; Holmstrom, K; Rajilic, M; Vaughan, E. E; et al. Colonic Microbiota
Signatures across Five Northern European Countries. Appl. Environ. Microbiol., 71(7):4153–
4155, 2005. doi:10.1128/AEM.71.7.4153-4155.2005. URL http://aem.asm.org/cgi/content/
abstract/71/7/4153.
Lee, Y. K; Menezes, J. S; Umesaki, Y; y Mazmanian, S. K. Proinflammatory T-cell respon-
ses to gut microbiota promote experimental autoimmune encephalomyelitis. Proceedings of
the National Academy of Sciences of the United States of America, 108 Suppl:4615–22, 2011.
ISSN 1091-6490. doi:10.1073/pnas.1000082107. URL http://www.pubmedcentral.nih.gov/
articlerender.fcgi?artid=3063590&tool=pmcentrez&rendertype=abstract.
Lennon, J. T y Jones, S. E. Microbial seed banks: the ecological and evolutionary implications
of dormancy. Nature reviews. Microbiology, 9(2):119–30, 2011. ISSN 1740-1534. doi:10.1038/
nrmicro2504. URL http://www.ncbi.nlm.nih.gov/pubmed/21233850.
Lenz, P y Søgaard-Andersen, L. Temporal and spatial oscillations in bacteria. Nature reviews.
Microbiology, 9(8):565–77, 2011. ISSN 1740-1534. doi:10.1038/nrmicro2612. URL http://www.
ncbi.nlm.nih.gov/pubmed/21760621.](https://image.slidesharecdn.com/0ed7e562-4fd2-456a-857e-89fd1b64755c-161012124743/85/tesis-156-320.jpg)