Descargado 28 veces









































































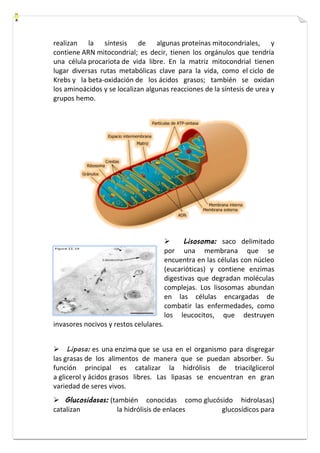













![cloroplastos, y el que se encuentra en el hialoplasma o en el retículo
endoplásmico de células eucariotas.
Cilios: son prolongaciones móviles localizadas en su superficie que
les permiten desplazarse en el entorno acuoso en que viven.
Flagelos: si tienen una longitud similar o superior a la de la propia
célula, apareciendo en este caso sólo uno.
Corpúsculo Basal: se localiza bajo la superficie celular
constituyendo la "raíz" del cilio o flagelo, tiene una estructura idéntica a
la de los centriolos, es decir, se trata de un cilindro hueco cuyas paredes
están formadas por nueve tripletes de microtúbulos (9×3).
Tallo o Axonema: el axonema, que se proyecta fuera de la célula,
está rodeado de membrana e interiormente presenta nueve pares de
microtúbulos periféricos y un par de microtúbulos centrales [(9×2)+2].
vacuolas](https://image.slidesharecdn.com/bibli-140829141535-phpapp01/85/BIBLIA-DE-LA-CELULA-96-320.jpg)


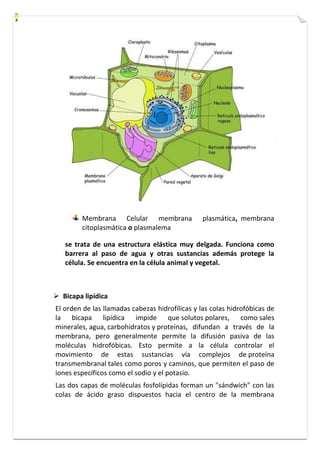


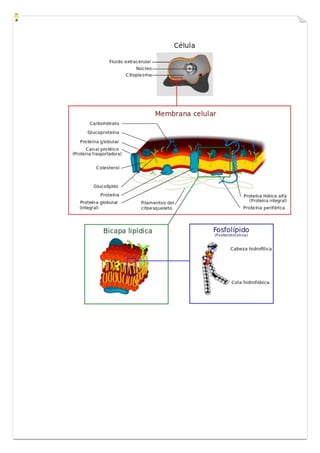
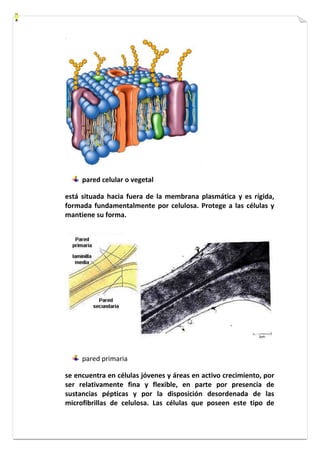







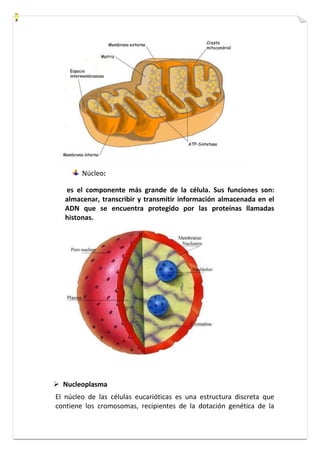





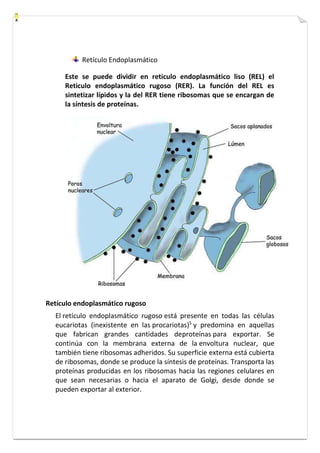
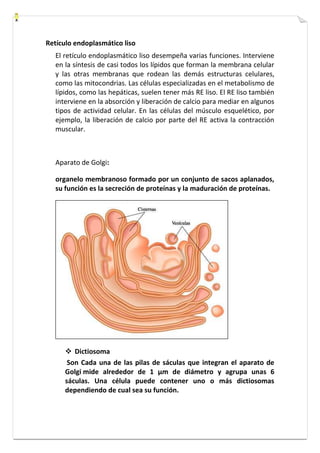


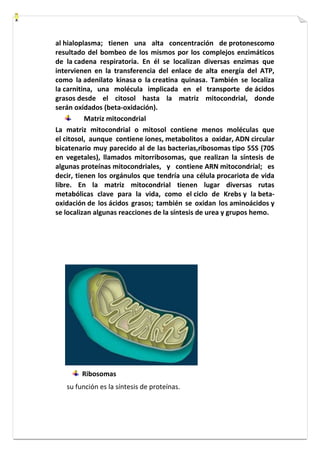




















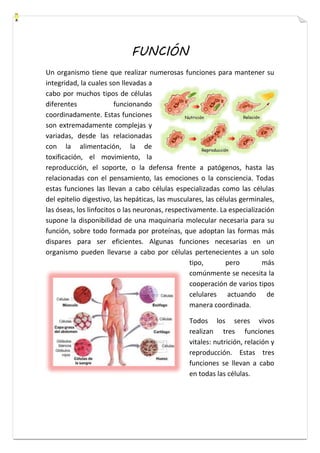
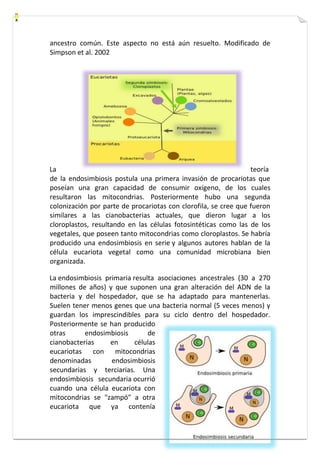
Este documento presenta una introducción a la biología celular. Explica que la célula es la unidad básica de los seres vivos y puede encontrarse de forma aislada o formando organismos pluricelulares. Describe las diferencias entre células procariotas y eucariotas y los principales compartimentos celulares como la membrana plasmática, núcleo, citosol y orgánulos. También habla sobre la diversidad en el tamaño y forma de las células en función de su función.














































































