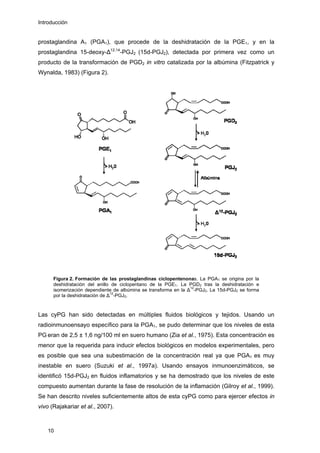
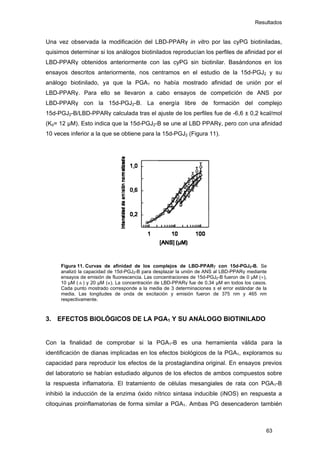
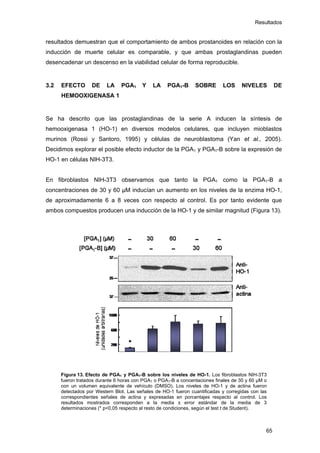
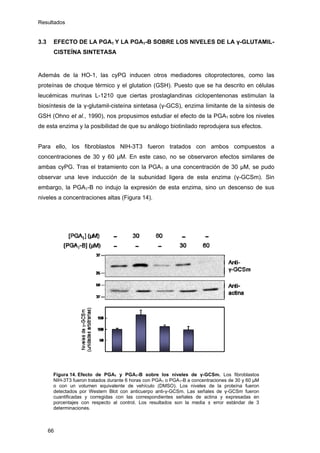
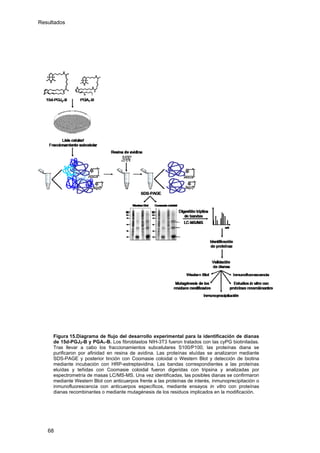
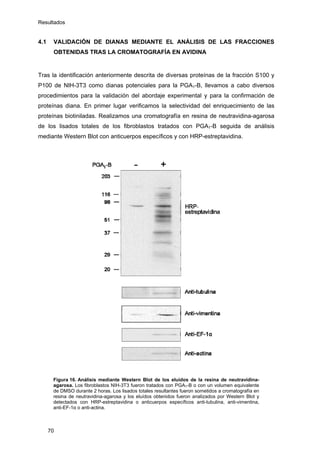
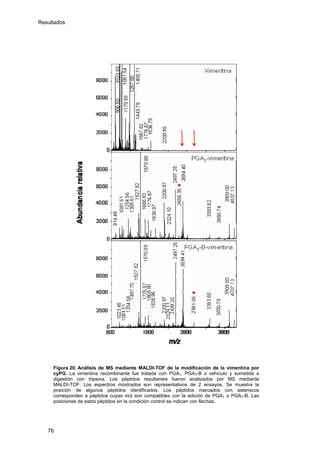
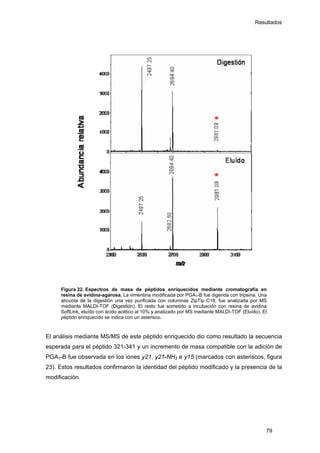
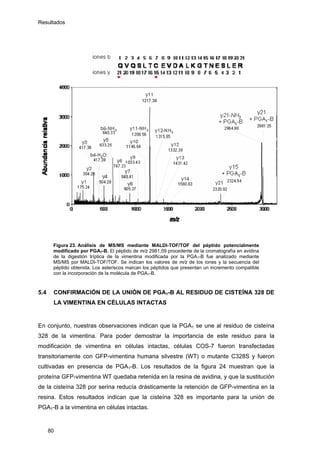
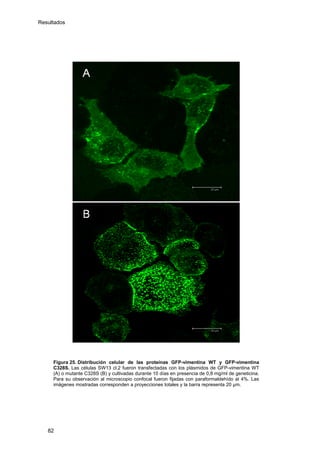
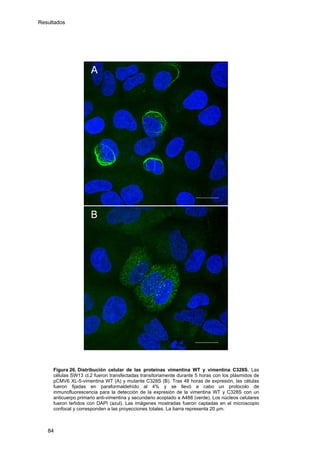
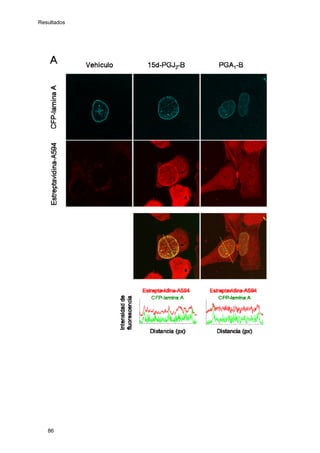
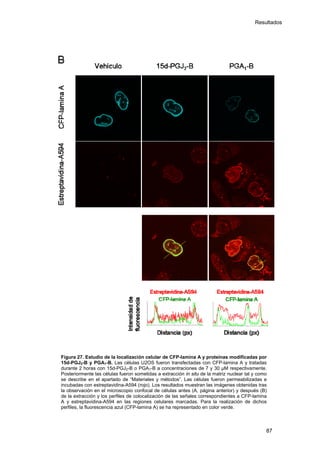
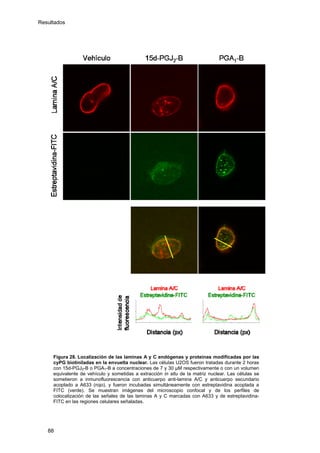
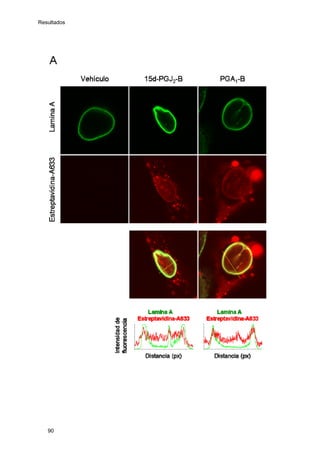
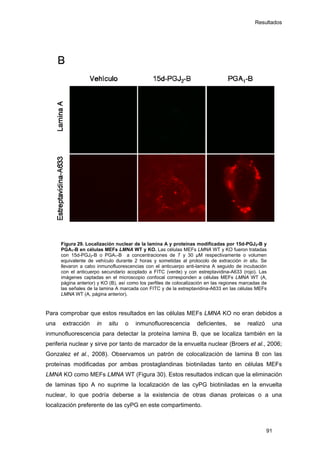
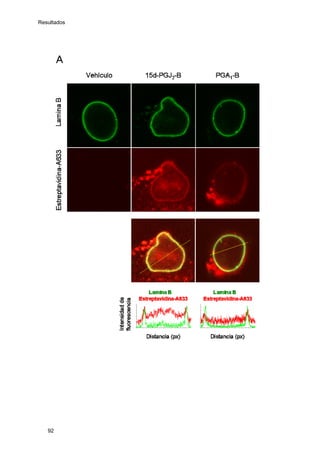
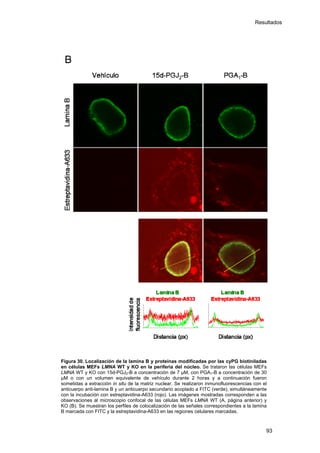
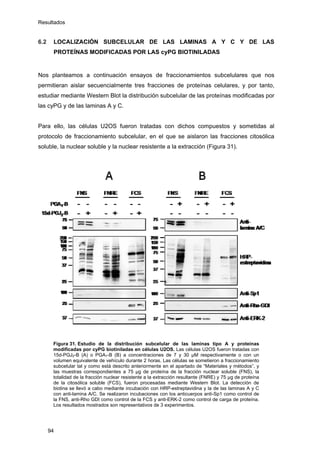
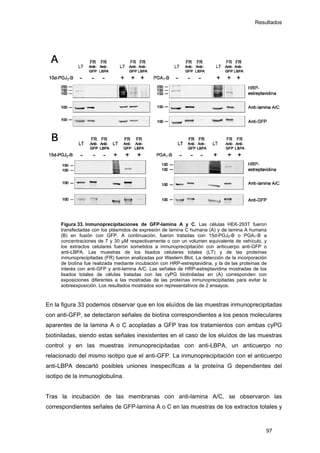
Este documento describe la memoria presentada por Beatriz Garzón Fernández para optar al título de Doctor en la Universidad Complutense de Madrid. La memoria se centra en la validación del uso del derivado biotinilado de la prostaglandina A1 para la identificación de sus dianas biológicas, así como en la caracterización de la modificación de proteínas de los filamentos intermedios como la vimentina y las laminas nucleares por prostaglandinas ciclopentenonas biotiniladas. El