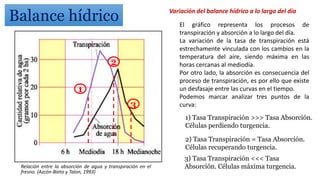
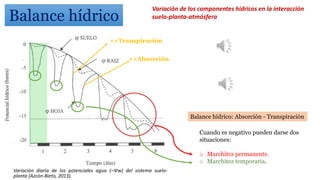
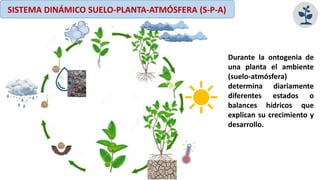
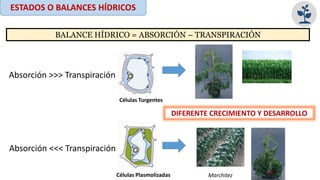
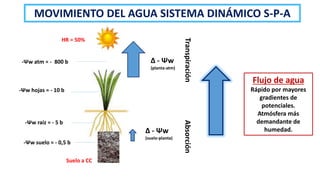
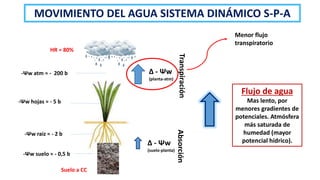
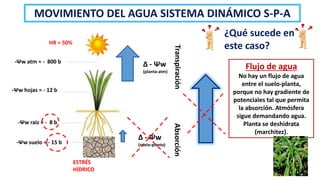
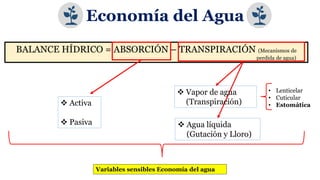
El documento analiza los mecanismos del movimiento de agua en el sistema suelo-planta-atmósfera y cómo esto determina los estados hídricos y el crecimiento de las plantas. Explica que la absorción y transpiración de agua dependen de los gradientes de potencial hídrico, y que la planta puede regular su balance hídrico mediante la apertura y cierre estomático, procesos influenciados por factores como la luz, el CO2 y el ácido abscísico.





![Conductividad
hidráulica
del
suelo
[m/h
.
MPa]
Potencia hídrico del suelo [MPa]
Capacidad Campo (CC)
Punto de marchitez
Permanente para la mayoría
de las especies (PMP)
Conductividad hidráulica para un suelo arcilloso en
función del potencial hídrico (Taiz & Seiger, 2015. Plan
Physiology and Development)
-Ψw
atmosférico
[bares]
Humedad relativa [%]
Potencial hídrico atmosférico en
función de la humedad relativa
ambiente.
VALORES DE POTENCIALES DE
REFERENCIA
PARA CONSIDERAR](https://image.slidesharecdn.com/claseeconomadelagua-parte1-240212030444-8c3fcc61/85/Clase-Economia-del-agua-Parte-1-ppsx-7-320.jpg)


![Tipos de Absorción
Activa (o movida por ósmosis)
• Baja demanda atmosférica (HR alta). Ritmo transpiratorio
lento.
• Presión xilemática positiva.
• Hay gasto de energía metabólica para acumular sustancias
osmóticamente activas (SOA) como azúcares, aminoácidos y/o
iones que disminuyen el potencial osmótico celular,
restableciendo así el gradiente de potenciales hídricos con el
suelo y se produzca la absorción.
-Ψw
(xilema)
[SOA] -Ψo -Ψw
v
Absorción
v
Ingresa agua](https://image.slidesharecdn.com/claseeconomadelagua-parte1-240212030444-8c3fcc61/85/Clase-Economia-del-agua-Parte-1-ppsx-15-320.jpg)


![Estimulada por:
Luz
CO2
CO2
↓ [CO2] estomática
FOTOSÍNTESIS H+
Bomba H+/ATPasa
Malato- + H+
K+
Cl-
Canales iónicos
Almidón de
reserva
↓- Ψo
↓- Ψw ↑- Ψw
INGRESO DE
AGUA
Apertura Estomática
Adaptado de Azcón-Bieto (2013)
Células
acompañantes](https://image.slidesharecdn.com/claseeconomadelagua-parte1-240212030444-8c3fcc61/85/Clase-Economia-del-agua-Parte-1-ppsx-20-320.jpg)

![Estimulada por:
Noche
CO2
CO2
↑ [CO2] estomática
FOTOSÍNTESIS
Bomba H+/ATPasa
↑[H2CO3] K+
Cl-
Canales iónicos
↑- Ψo
↑- Ψw ↓- Ψw
CO2
CO2
CO2
↓ pH
SALIDA DE
AGUA
Cierre Estomático
↓ [Malato- + H+ ]
Adaptado de Azcón-Bieto (2013) y Bidwell (1993)
Células
acompañantes](https://image.slidesharecdn.com/claseeconomadelagua-parte1-240212030444-8c3fcc61/85/Clase-Economia-del-agua-Parte-1-ppsx-23-320.jpg)

![Restricción de agua Agua disponible
Tiempo [días]
Resistencia
estomática
[s/cm]
Potencial
hídrico
hoja
[Mpa]
ABA
[ng/cm
2
hoja]
El potencial hídrico disminuye
conforme el suelo se seca
Resistencia
estomática
Contenido
ABA
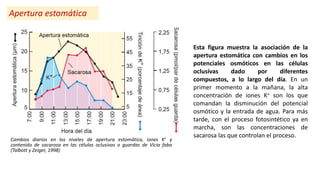
Cambios en el potencial hídrico, en la resistencia
estomática y en la concentración de ABA en maíz, en
respuesta al estrés hídrico (Bearsell y Cohen, 1975).
En la figura se puede ver claramente como
aumenta la concentración de ABA en respuesta a
una situación de estrés hídrico, y la relación del
aumento de ABA con el aumento resistencia
estomática, variable vinculada con el cierre
estomático.
La resistencia estomática disminuye como
consecuencia de menores valores de ABA cuando
se restituye la disponibilidad hídrica.
Esto demuestra que el cierre estomático regulado
por ABA es una estrategia adaptativa de las
plantas a condiciones de estrés ambiental.
Acción del Ácido Abscisico
(ABA)
Existe otro mecanismo fisiológico por el cual las plantas pueden
regular el cierre de sus estomas, ANTE CONDICIONES DE ESTRES:](https://image.slidesharecdn.com/claseeconomadelagua-parte1-240212030444-8c3fcc61/85/Clase-Economia-del-agua-Parte-1-ppsx-25-320.jpg)
![Estimulada por:
ESTRÉS HÍDRICO
Bomba H+/ATPasa
K+
Cl-
Canales iónicos
↑- Ψo
↑- Ψw ↓- Ψw
ABA
Luz
CO2
CO2
↓ [CO2] estomática
↑ pH
SALIDA DE
AGUA
Cierre Estomático (ABA)
↑ [Ca+]
Receptor
Adaptado de Azcón-Bieto (2013) y García-Breijo (2001)
Células
acompañantes](https://image.slidesharecdn.com/claseeconomadelagua-parte1-240212030444-8c3fcc61/85/Clase-Economia-del-agua-Parte-1-ppsx-26-320.jpg)