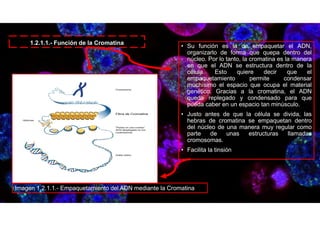
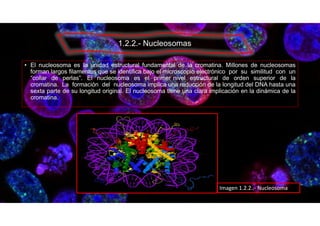
Este documento describe la estructura y función de los organelos de las células. Explica que el núcleo alberga el ADN de la célula y contiene poros nucleares que permiten el transporte entre el núcleo y el citoplasma. También describe otros componentes del material genético como la cromatina, nucleosomas y cromosomas, que ayudan a empaquetar y organizar el ADN. El objetivo es conocer los diferentes organelos de las células animales, vegetales y bacterias, y comprender sus funciones.

































































































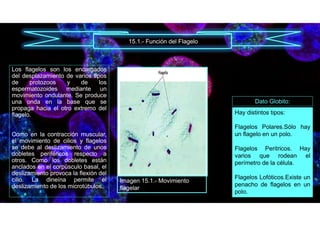











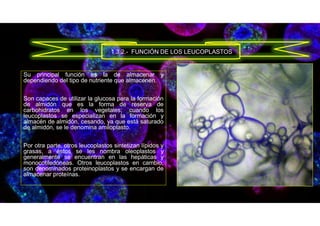
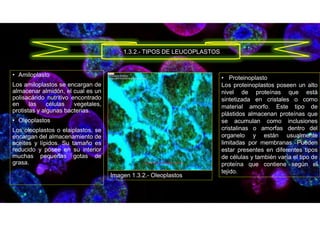




























![• E. (2019, 6 noviembre). Función de los Cromoplastos3.5 (70%) 2 voto[s] Un cromoplasto es un
plasto Leer más. Funcion.info. https://www.funcion.info/cromoplastos/
• Perez, M. (2019, 26 noviembre). Leucoplastos: características, tipos y funciones. Lifeder.
https://www.lifeder.com/leucoplastos/
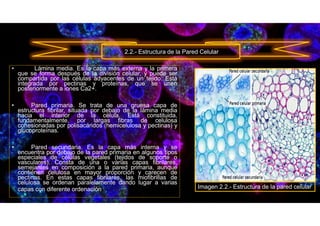
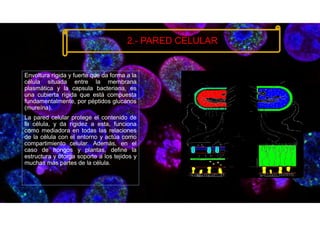
• 2.3.2. Pared celular en células vegetales. (s. f.). Biologia sur.
https://www.biologiasur.org/index.php/145-apuntes-de-biologia/procariotica-y-eucariotica/334-2-3-2-
pared-celular-en-celulas-vegetales
• Gelambi, M. (2019b, agosto 24). Nucleoide: características, estructura, composición, funciones.
Lifeder. https://www.lifeder.com/nucleoide/
• Citoplasma y su contenido. (s. f.). Citoplasma. Recuperado 17 de octubre de 2020, de
https://ugr.es/%7Eeianez/Microbiologia/07citoplasma.htm
• Gómez, C. F. L. (2019, 11 febrero). Mesosomas: características, tipos y funciones. Lifeder.
https://www.lifeder.com/mesosomas/
• http://www.higiene.edu.uy/cefa/2008/MorfologiayEstructuraBacteriana.pdf
• https://biologia-geologia.com/biologia2](https://image.slidesharecdn.com/atlasdelacelula-201019195926/85/Atlas-de-la-celula-152-320.jpg)
























































