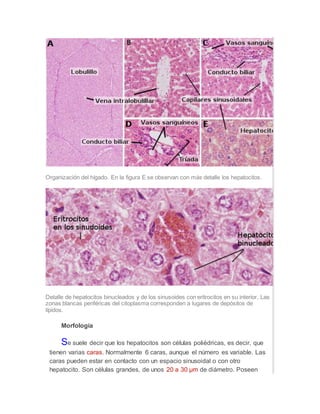
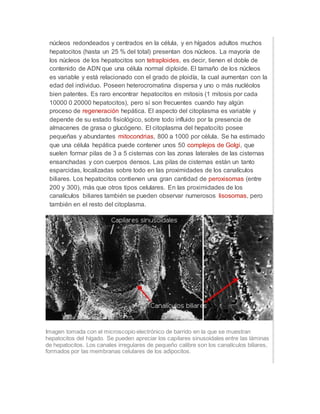
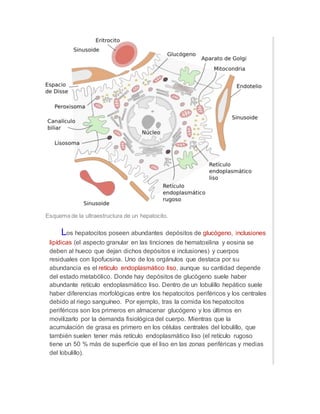
Los hepatocitos son las células principales del hígado y desempeñan un papel clave en el metabolismo. Contienen numerosos orgánulos como mitocondrias, retículo endoplásmico y peroxisomas que les permiten sintetizar moléculas, almacenar glucógeno y grasas, y desintoxicar compuestos. Los hepatocitos forman los canalículos biliares que secretan la bilis y están polarizados para realizar eficientemente sus funciones metabólicas y detoxificación.