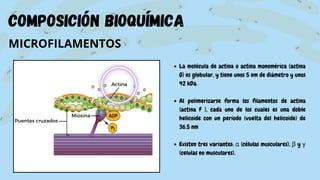
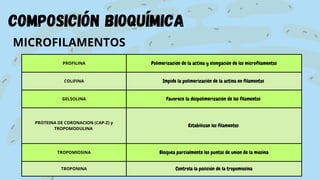
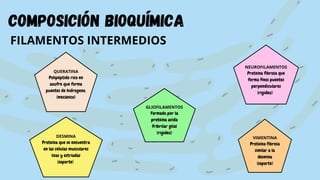
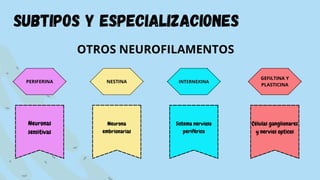
El documento proporciona información sobre el citoesqueleto, la red tridimensional de filamentos proteicos dentro de la célula que le da forma y estructura. Describe los tres tipos principales de filamentos - microfilamentos, filamentos intermedios y microtúbulos - así como sus funciones en dar soporte estructural, permitir la movilidad celular y transporte intracelular. También discute las diferencias en la composición y función del citoesqueleto entre células animales, vegetales, eucariotas y procariotas.