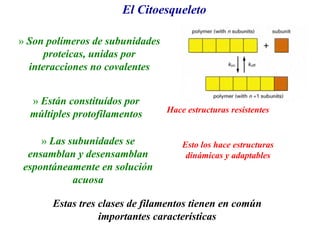
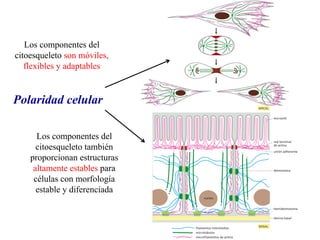
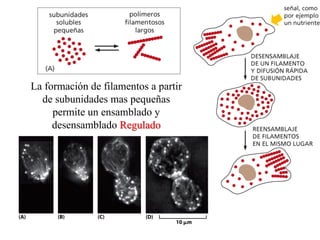
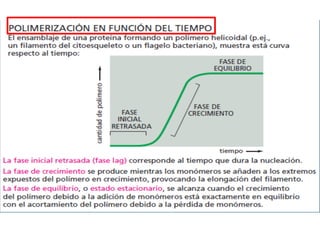
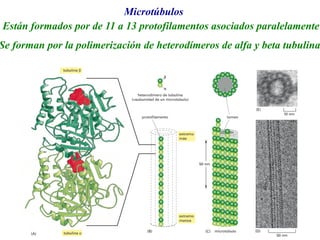
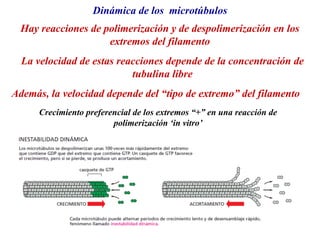
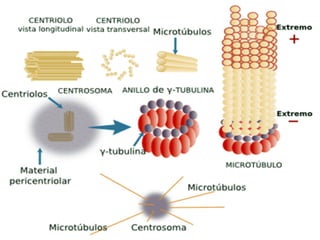
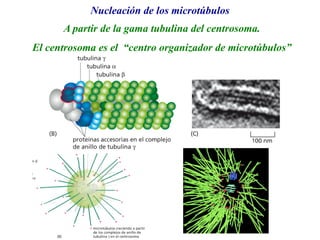
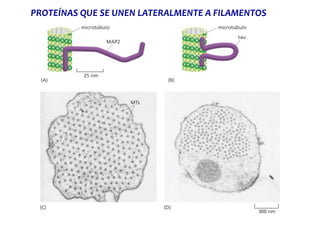
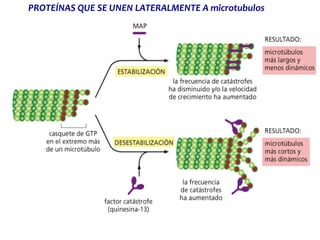
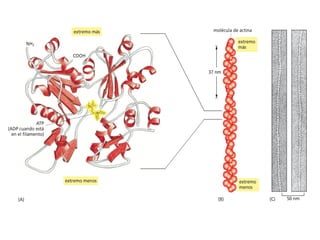
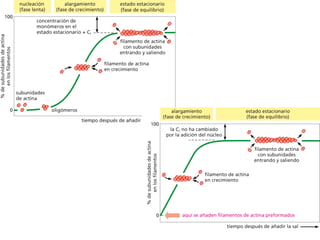
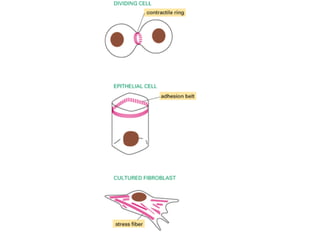
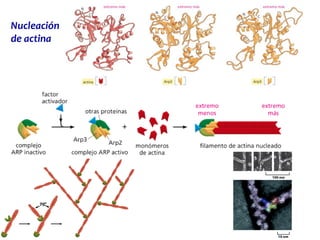
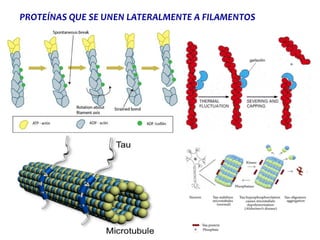
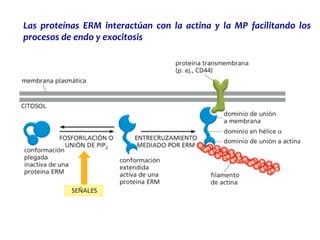
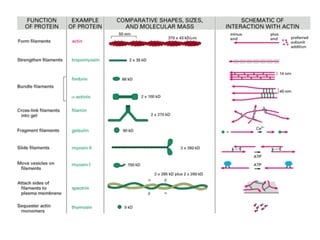
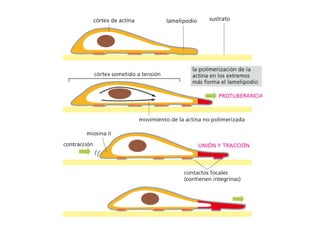
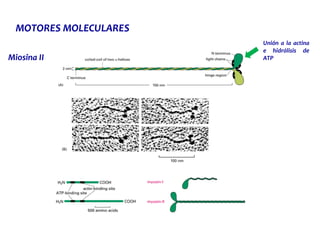
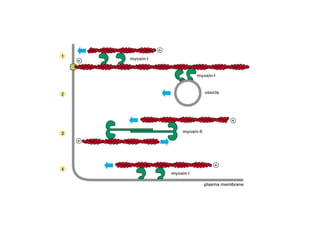
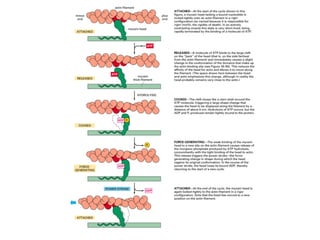
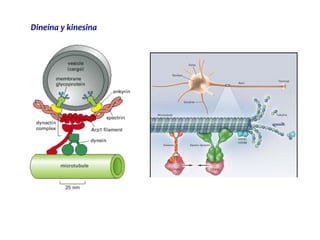
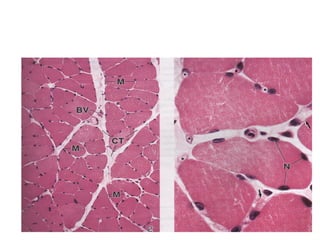
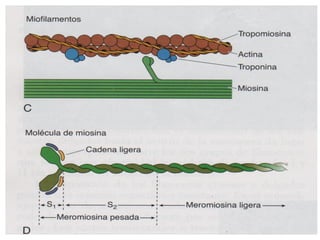
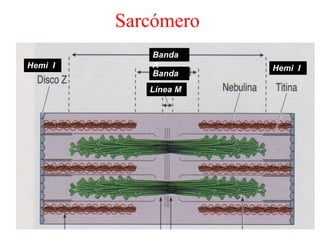
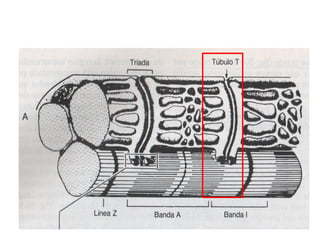
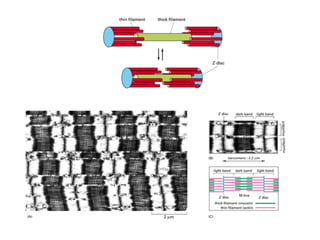
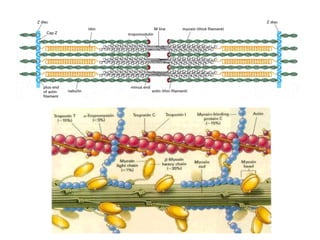
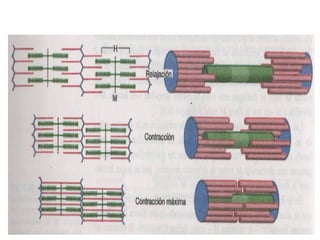
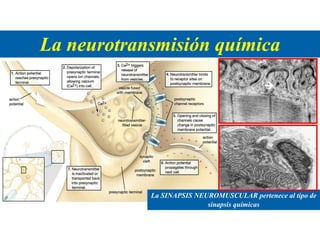
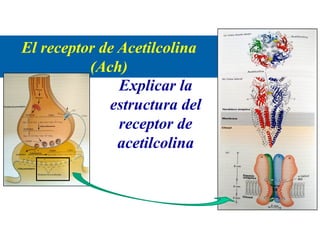
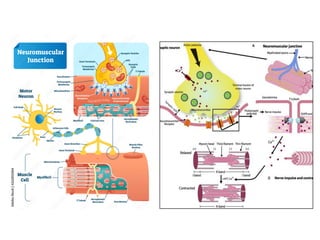
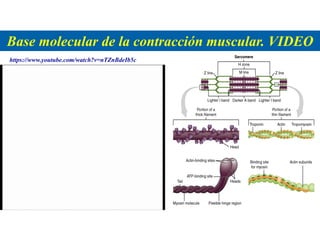
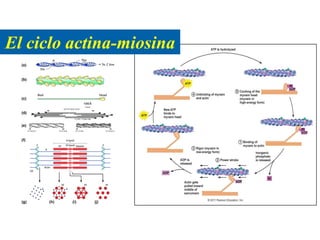
El documento resume las características principales de los tres tipos de filamentos que componen el citoesqueleto: filamentos intermedios, microtúbulos y filamentos de actina. Explica que cada tipo de filamento está formado por la polimerización de proteínas específicas y desempeña funciones estructurales y funcionales únicas en la célula. Además, los tres tipos de filamentos interactúan entre sí a través de proteínas asociadas para permitir el movimiento celular y el transporte intracelular.