Descargado 32 veces









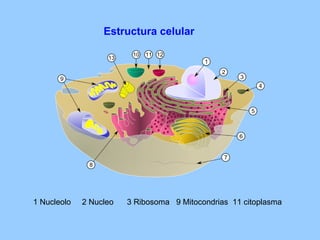

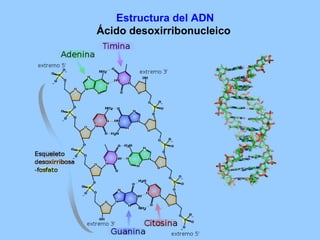
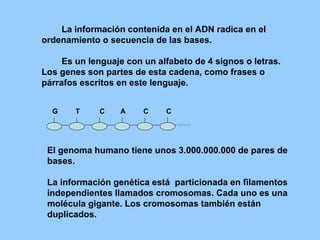
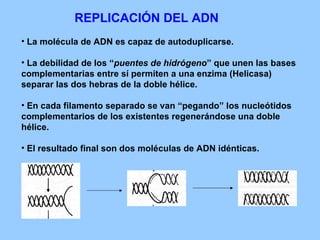
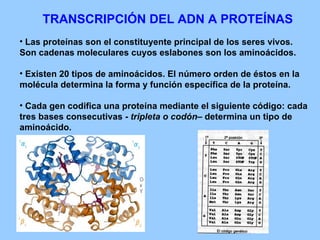

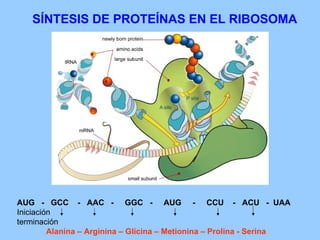
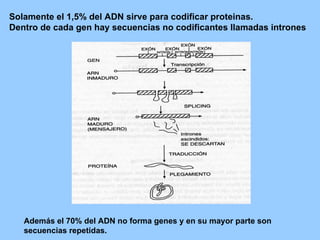
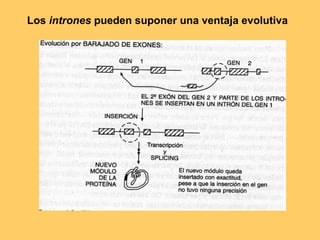

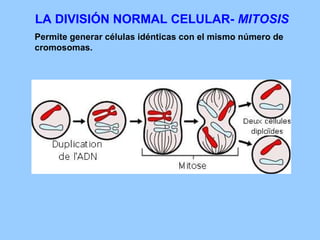
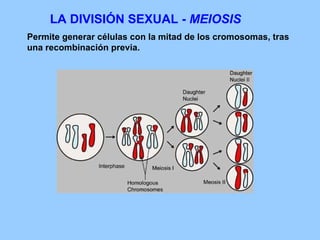


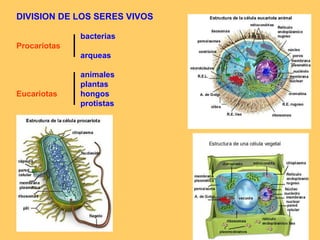


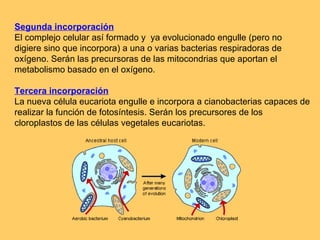
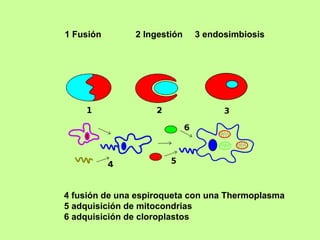
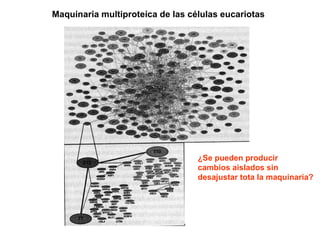



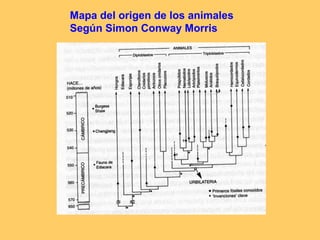
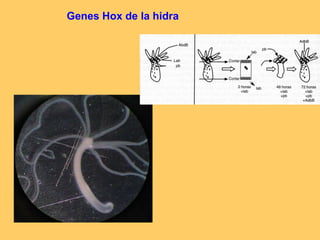

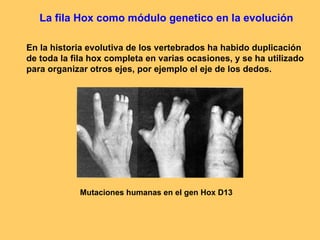
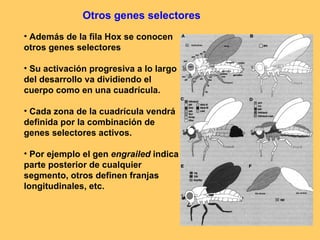


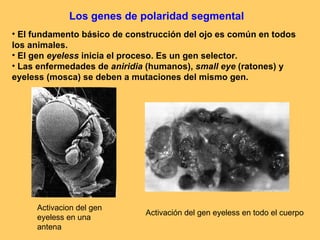




El documento resume la evolución de las ideas sobre la evolución biológica desde Darwin hasta la actualidad. Comienza con las ideas de Darwin y Wallace sobre la selección natural. Luego describe teorías posteriores como el neolamarckismo, ortogénesis y saltacionismo. Explica la síntesis evolutiva moderna y teorías posteriores como el equilibrio puntuado. Finalmente, aborda temas como la endosimbiosis, los genes Hox y la evolución del ojo.



























































































