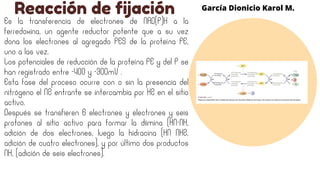
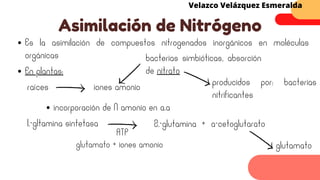
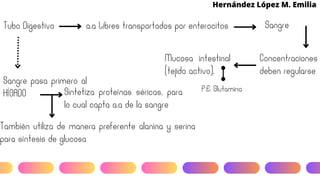
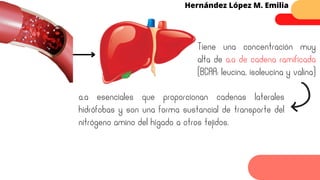
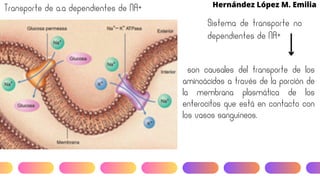
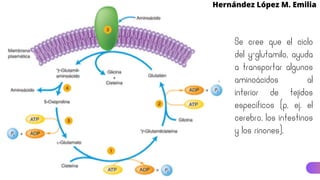
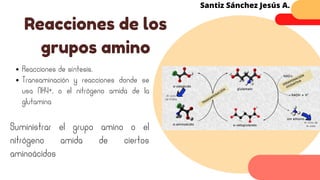
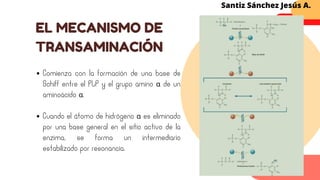
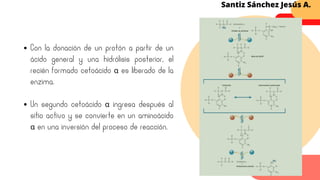
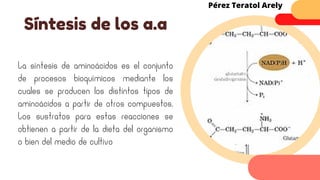
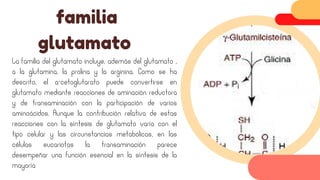
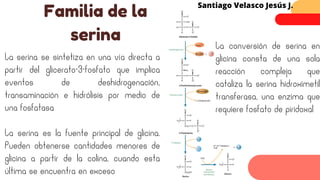
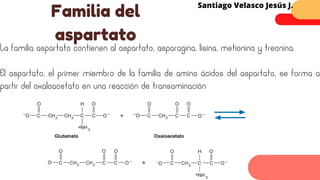
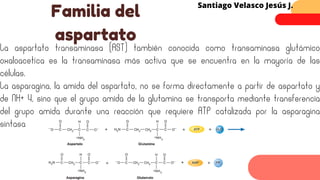
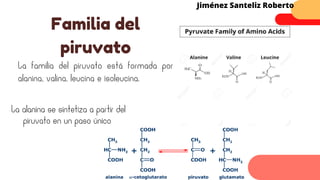
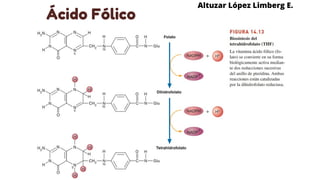
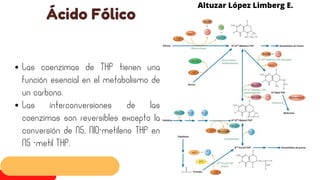
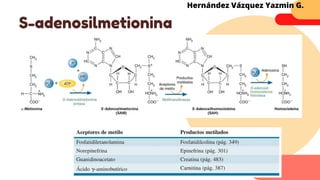
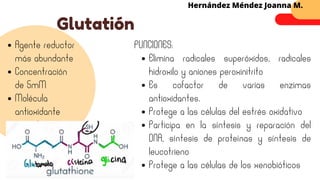
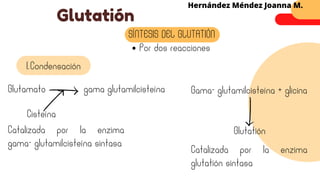
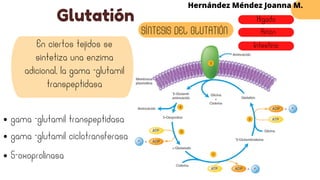
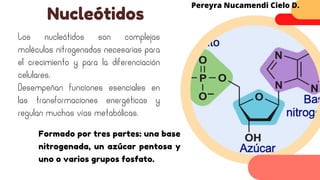
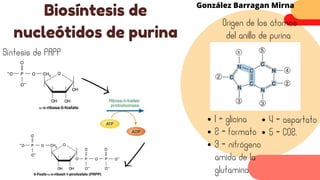
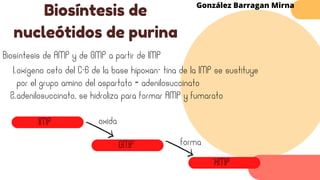
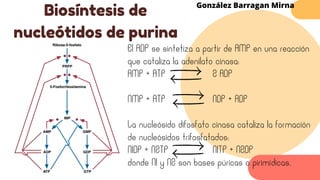
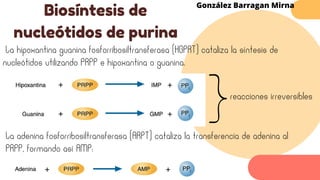
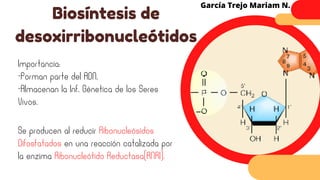
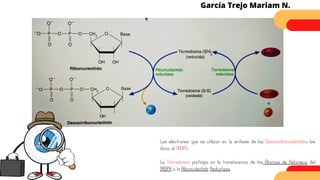
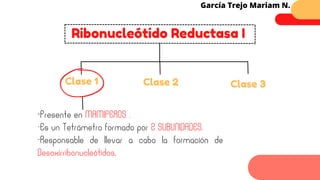
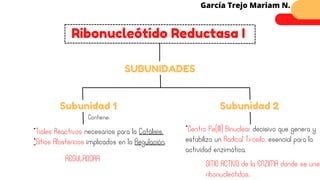
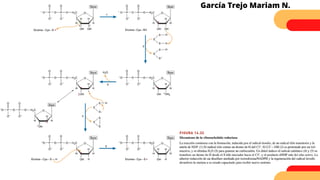
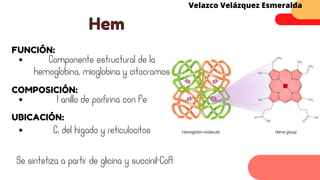
Este documento describe el metabolismo del nitrógeno en plantas y animales. Se discuten los procesos de fijación de nitrógeno, asimilación de nitrógeno, síntesis de aminoácidos y metabolismo de aminoácidos. También se describen las familias de aminoácidos como glutamato, serina, aspartato, piruvato y aromáticos, así como sus rutas biosintéticas. Finalmente, se mencionan otros metabolitos relacionados con el nitrógeno como glutatión, S-adenosilmetionina y ácido f