Descargado 10 veces







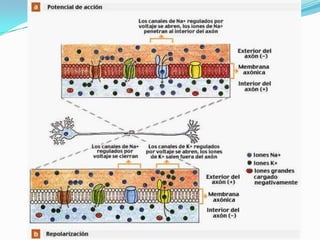



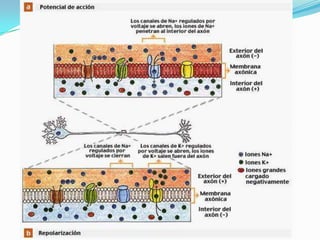



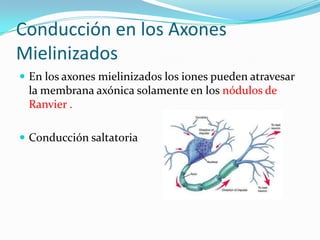













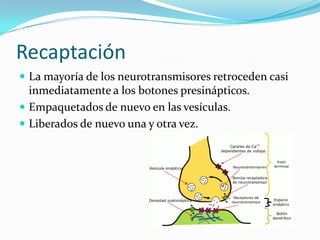




























El documento resume las bases iónicas y moleculares de la transmisión sináptica. Explica cómo los potenciales de acción se conducen a lo largo del axón mediante canales iónicos activados por voltaje y cómo diferentes neurotransmisores como la acetilcolina, el glutamato y las aminas biógenas se sintetizan, almacenan, liberan y degradan en las sinapsis para transmitir mensajes entre neuronas.















































