PROPIEDADES BIOQUÍMICAS DEL
GLÓBULOROJO
Hematología e Inmunología - UC13 2023
Dr. Nicolás Campolo
Departamento de Bioquímica
Facultad de Medicina - UdelaR
Glóbulo rojo (eritrocito)
oTipo celular más numeroso en la sangre
o Ocupan aproximadamente un 40% del
volumen sanguíneo total (hematocrito)
o Funciones principales: transporte de
gases (O2 y CO2) y regulación del pH del
plasma
o Estructura simple; un 95% de la proteína
intracelular es hemoglobina (Hb)
o Pese a esto, NO es una célula inerte desde
el punto de vista metabólico
4.
Eritropoyesis
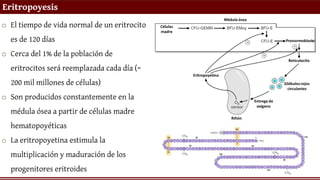
o El tiempode vida normal de un eritrocito
es de 120 días
o Cerca del 1% de la población de
eritrocitos será reemplazada cada día (~
200 mil millones de células)
o Son producidos constantemente en la
médula ósea a partir de células madre
hematopoyéticas
o La eritropoyetina estimula la
multiplicación y maduración de los
progenitores eritroides
5.
Destrucción de losglóbulos rojos
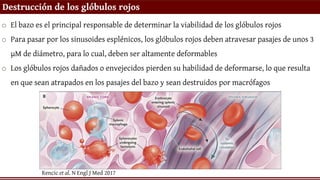
o El bazo es el principal responsable de determinar la viabilidad de los glóbulos rojos
o Para pasar por los sinusoides esplénicos, los glóbulos rojos deben atravesar pasajes de unos 3
µM de diámetro, para lo cual, deben ser altamente deformables
o Los glóbulos rojos dañados o envejecidos pierden su habilidad de deformarse, lo que resulta
en que sean atrapados en los pasajes del bazo y sean destruidos por macrófagos
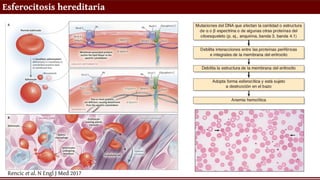
Rencic et al. N Engl J Med 2017
Morfología celular
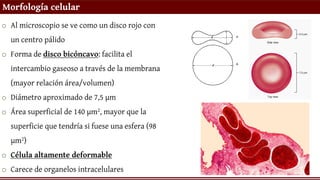
o Almicroscopio se ve como un disco rojo con
un centro pálido
o Forma de disco bicóncavo: facilita el
intercambio gaseoso a través de la membrana
(mayor relación área/volumen)
o Diámetro aproximado de 7,5 µm
o Área superficial de 140 µm2, mayor que la
superficie que tendría si fuese una esfera (98
µm2)
o Célula altamente deformable
o Carece de organelos intracelulares
8.
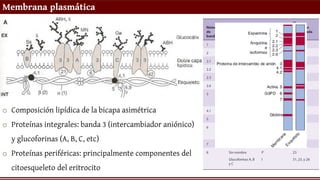
Membrana plasmática
o Composiciónlipídica de la bicapa asimétrica
o Proteínas integrales: banda 3 (intercambiador aniónico)
y glucoforinas (A, B, C, etc)
o Proteínas periféricas: principalmente componentes del
citoesqueleto del eritrocito
9.
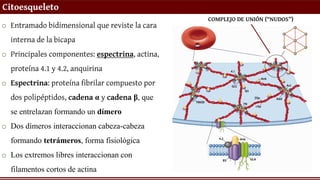
Citoesqueleto
o Entramado bidimensionalque reviste la cara
interna de la bicapa
o Principales componentes: espectrina, actina,
proteína 4.1 y 4.2, anquirina
o Espectrina: proteína fibrilar compuesto por
dos polipéptidos, cadena α y cadena β, que
se entrelazan formando un dímero
o Dos dímeros interaccionan cabeza-cabeza
formando tetrámeros, forma fisiológica
o Los extremos libres interaccionan con
filamentos cortos de actina
COMPLEJO DE UNIÓN (“NUDOS”)
10.
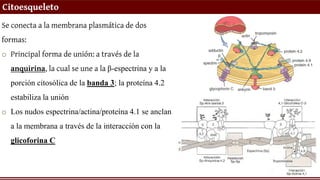
Citoesqueleto
Se conecta ala membrana plasmática de dos
formas:
o Principal forma de unión: a través de la
anquirina, la cual se une a la β-espectrina y a la
porción citosólica de la banda 3; la proteína 4.2
estabiliza la unión
o Los nudos espectrina/actina/proteína 4.1 se anclan
a la membrana a través de la interacción con la
glicoforina C
11.
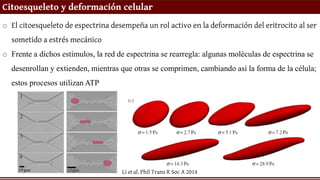
Citoesqueleto y deformacióncelular
o El citoesqueleto de espectrina desempeña un rol activo en la deformación del eritrocito al ser
sometido a estrés mecánico
o Frente a dichos estímulos, la red de espectrina se rearregla: algunas moléculas de espectrina se
desenrollan y extienden, mientras que otras se comprimen, cambiando así la forma de la célula;
estos procesos utilizan ATP
Li et al. Phil Trans R Soc A 2014
12.
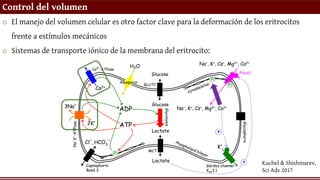
Control del volumen
oEl manejo del volumen celular es otro factor clave para la deformación de los eritrocitos
frente a estímulos mecánicos
o Sistemas de transporte iónico de la membrana del eritrocito:
Kuchel & Shishmarev,
Sci Adv 2017
13.
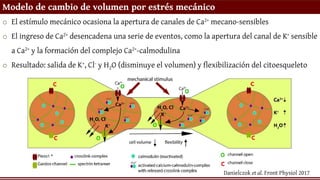
Modelo de cambiode volumen por estrés mecánico
o El estímulo mecánico ocasiona la apertura de canales de Ca2+ mecano-sensibles
o El ingreso de Ca2+ desencadena una serie de eventos, como la apertura del canal de K+ sensible
a Ca2+ y la formación del complejo Ca2+-calmodulina
o Resultado: salida de K+, Cl- y H2O (disminuye el volumen) y flexibilización del citoesqueleto
Danielczok et al. Front Physiol 2017
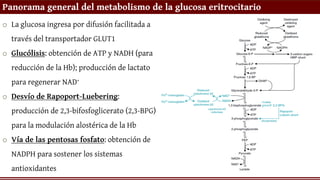
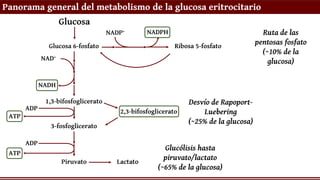
Panorama general delmetabolismo de la glucosa eritrocitario
o La glucosa ingresa por difusión facilitada a
través del transportador GLUT1
o Glucólisis: obtención de ATP y NADH (para
reducción de la Hb); producción de lactato
para regenerar NAD+
o Desvío de Rapoport-Luebering:
producción de 2,3-bifosfoglicerato (2,3-BPG)
para la modulación alostérica de la Hb
o Vía de las pentosas fosfato: obtención de
NADPH para sostener los sistemas
antioxidantes
17.
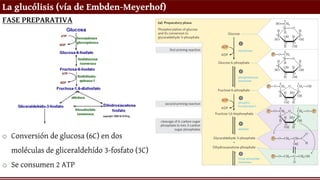
La glucólisis (víade Embden-Meyerhof)
FASE PREPARATIVA
o Conversión de glucosa (6C) en dos
moléculas de gliceraldehído 3-fosfato (3C)
o Se consumen 2 ATP
18.
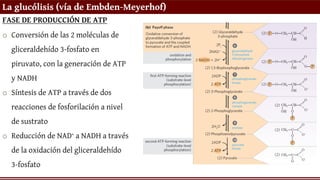
La glucólisis (víade Embden-Meyerhof)
FASE DE PRODUCCIÓN DE ATP
o Conversión de las 2 moléculas de
gliceraldehído 3-fosfato en
piruvato, con la generación de ATP
y NADH
o Síntesis de ATP a través de dos
reacciones de fosforilación a nivel
de sustrato
o Reducción de NAD+ a NADH a través
de la oxidación del gliceraldehído
3-fosfato
19.
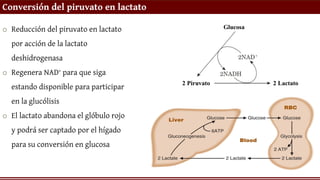
Conversión del piruvatoen lactato
o Reducción del piruvato en lactato
por acción de la lactato
deshidrogenasa
o Regenera NAD+ para que siga
estando disponible para participar
en la glucólisis
o El lactato abandona el glóbulo rojo
y podrá ser captado por el hígado
para su conversión en glucosa
20.
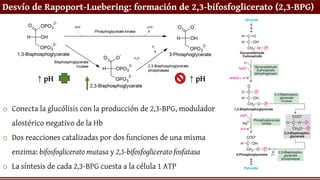
Desvío de Rapoport-Luebering:formación de 2,3-bifosfoglicerato (2,3-BPG)
o Conecta la glucólisis con la producción de 2,3-BPG, modulador
alostérico negativo de la Hb
o Dos reacciones catalizadas por dos funciones de una misma
enzima: bifosfoglicerato mutasa y 2,3-bifosfoglicerato fosfatasa
o La síntesis de cada 2,3-BPG cuesta a la célula 1 ATP
↑ pH ↑ pH
21.
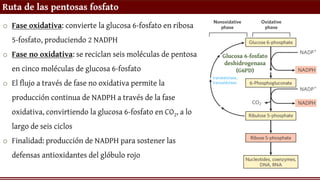
Ruta de laspentosas fosfato
o Fase oxidativa: convierte la glucosa 6-fosfato en ribosa
5-fosfato, produciendo 2 NADPH
o Fase no oxidativa: se reciclan seis moléculas de pentosa
en cinco moléculas de glucosa 6-fosfato
o El flujo a través de fase no oxidativa permite la
producción continua de NADPH a través de la fase
oxidativa, convirtiendo la glucosa 6-fosfato en CO2, a lo
largo de seis ciclos
o Finalidad: producción de NADPH para sostener las
defensas antioxidantes del glóbulo rojo
Glucosa 6-fosfato
deshidrogenasa
(G6PD)
22.
Panorama general delmetabolismo de la glucosa eritrocitario
Glucosa
Glucosa 6-fosfato Ribosa 5-fosfato
Ruta de las
pentosas fosfato
(~10% de la
glucosa)
NADP+ NADPH
1,3-bifosfoglicerato
3-fosfoglicerato
2,3-bifosfoglicerato
Desvío de Rapoport-
Luebering
(~25% de la glucosa)
Piruvato Lactato
Glucólisis hasta
piruvato/lactato
(~65% de la glucosa)
NAD+
NADH
ADP
ATP
ADP
ATP
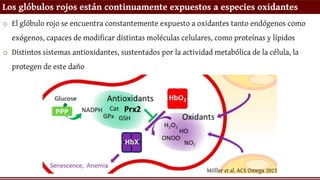
Los glóbulos rojosestán continuamente expuestos a especies oxidantes
Möller et al. ACS Omega 2023
o El glóbulo rojo se encuentra constantemente expuesto a oxidantes tanto endógenos como
exógenos, capaces de modificar distintas moléculas celulares, como proteínas y lípidos
o Distintos sistemas antioxidantes, sustentados por la actividad metabólica de la célula, la
protegen de este daño
25.
Autooxidación de laHb: generación endógena de oxidantes
o Consecuencias: formación de MetHb (no transporta O2) y de una especie oxidante, el O2
•-
Hb(Fe2+)-O2 → Hb(Fe3+) + O2
•-
Oxihemoglobina Metahemoglobina Radical anión superóxido
o Efectos deletéreos de la formación de O2
•-:
• Puede oxidar a la Hb
• Formación de oxidantes secundarios más fuertes
O2
•- + O2
•- + 2H+ → H2O2 + O2
Peróxido de hidrógeno
o Dentro del eritrocito, la dismutación del O2
•- a H2O2 es catalizada por la enzima superóxido
dismutasa (SOD)
26.
Autooxidación de laHb: generación endógena de oxidantes
o Reacción del H2O2 con el hemo: generación de MetHb
Hb(Fe2+)-O2 + H2O2 → Hb(Fe4+=O) + H2O + O2
o Reacción del H2O2 con Fe2+ lábil: formación de •OH
(reacción de Fenton)
Hb(Fe4+=O) + H2O2 → Hb(Fe3+) + H2O + O2
•-
Ferrilhemoglobina
Metahemoglobina
o Reacción del H2O2 con las globinas: desnaturalización de la Hb, formación de precipitados
(cuerpos de Heinz)
Fe2+X + H2O2 → Fe3+X + •OH + OH-
Radical hidroxilo
27.
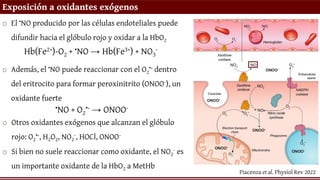
Exposición a oxidantesexógenos
o El •NO producido por las células endoteliales puede
difundir hacia el glóbulo rojo y oxidar a la HbO2
Hb(Fe2+)-O2 + •NO → Hb(Fe3+) + NO3
-
Piacenza et al. Physiol Rev 2022
o Además, el •NO puede reaccionar con el O2
•- dentro
del eritrocito para formar peroxinitrito (ONOO-), un
oxidante fuerte
•NO + O2
•- → ONOO-
o Otros oxidantes exógenos que alcanzan el glóbulo
rojo: O2
•-, H2O2, NO2
-, HOCl, ONOO-
o Si bien no suele reaccionar como oxidante, el NO2
- es
un importante oxidante de la HbO2 a MetHb
28.
Defensas antioxidantes delglóbulo rojo
o Detoxificación del O2
•-: superóxido dismutasa de cobre/zinc (SOD1)
o Detoxificación del H2O2: tres sistemas enzimáticos principales
1. Peroxirredoxina (Prx)/ tiorredoxina (Trx)/ tiorredoxina reductasa (TR)
2. Glutatión peroxidasa (GPx)/ glutatión (GSH)/ glutatión reductasa (GR)
3. Catalasa
O2
•- + O2
•- + 2H+
Hb(Fe2+)-O2
Hb(Fe3+)
H2O2 + O2
SOD
o La actividad de los sistemas redutores Prx/Trx/TR y GPx/GSH/GR se sostiene en base al
poder reductor del NADPH: dependen de la actividad metabólica (vía pentosas fosfato)
o Antioxidantes de bajo peso molecular: glutatión, ácido ascórbico (vitamina C), α-tocoferol
(vitamina E) y ácido úrico
29.
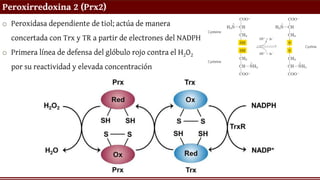
Peroxirredoxina 2 (Prx2)
oPeroxidasa dependiente de tiol; actúa de manera
concertada con Trx y TR a partir de electrones del NADPH
o Primera línea de defensa del glóbulo rojo contra el H2O2
por su reactividad y elevada concentración
30.
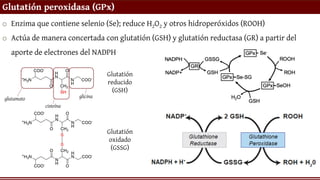
Glutatión peroxidasa (GPx)
oEnzima que contiene selenio (Se); reduce H2O2 y otros hidroperóxidos (ROOH)
o Actúa de manera concertada con glutatión (GSH) y glutatión reductasa (GR) a partir del
aporte de electrones del NADPH
Glutatión
reducido
(GSH)
Glutatión
oxidado
(GSSG)
glutamato
cisteína
glicina
31.
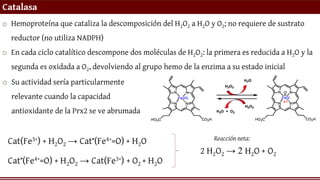
Catalasa
o Hemoproteína quecataliza la descomposición del H2O2 a H2O y O2; no requiere de sustrato
reductor (no utiliza NADPH)
o En cada ciclo catalítico descompone dos moléculas de H2O2: la primera es reducida a H2O y la
segunda es oxidada a O2, devolviendo al grupo hemo de la enzima a su estado inicial
Cat(Fe3+) + H2O2 → Cat•(Fe4+=O) + H2O
Cat•(Fe4+=O) + H2O2 → Cat(Fe3+) + O2 + H2O
2 H2O2 → 2 H2O + O2
Reacción neta:
o Su actividad sería particularmente
relevante cuando la capacidad
antioxidante de la Prx2 se ve abrumada
32.
Sistemas de reparación:metahemoglobina reductasa
o Entre un 0,5-3% de la HbO2 se autooxida diariamente; no obstante, menos del 1% de la Hb se
encuentra como MetHb en condiciones normales
o Esto se debe a que el glóbulo rojo cuenta con un sistema enzimático responsable de catalizar
la reducción de la MetHb: la metahemoglobina reductasa (NADH-citocromo b5 metHb
reductasa), que usa los electrones del NADH (¡no NADPH!) para reducir el Fe3+
33.
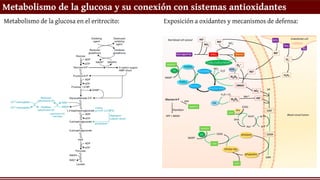
Metabolismo de laglucosa y su conexión con sistemas antioxidantes
Metabolismo de la glucosa en el eritrocito: Exposición a oxidantes y mecanismos de defensa:
Bibliografía
Harper Bioquímica Ilustrada(Capítulo 52) - Murray, R.K., Bender, D.A., Botham, K.M., Kennelly, P.J., Rodwell,
V.W. & Weil, P.A. 29ª Edición, Ed. McGraw-Hill, 2013.
Mark’s Basic Medical Biochemistry (Capítulo 44) - Smith, C.M., Marks, A.D., & Lieberman, M.A. 2ª Edición, Ed.
Lippincott Williams & Wilkins, 2005.
Hematología clínica (Capítulo 4) - Sans-Sabrafen, J., Besses Raebel, C. & Vives Corrons, J.L. 5ª Edición, Ed. Elsevier
España, 2006.
Möller, M., Orrico, F., Villar, S.F., López, A.C., Silva, N., Donzé, M., Thomson, L. & Denicola, A. “Oxidants and
Antioxidants in the Redox Biochemistry of Human Red Blood Cells”, ACS Omega, 8, 147-168, 2023.
Huisjes, R., Bogdanova, A., van Solinge, W.W., Schiffelers, R.M., Kaestner, L. & van Wijk, R. “Squeezing for Life -
Properties of Red Blood Cell Deformability”, Frontiers in Physiology, 9:656, 2018