Descargado 29 veces










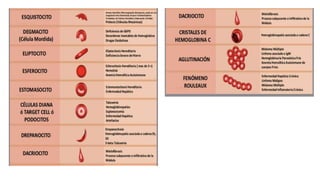








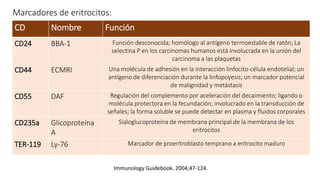








El documento describe el proceso de eritropoyesis, donde se generan eritrocitos a partir de células madre hematopoyéticas a través de diferentes etapas de diferenciación. Destaca la estructura y función de los eritrocitos, incluyendo la hemoglobina y su papel en el transporte de oxígeno y dióxido de carbono. Además, se analizan las variaciones en la eritropoyesis entre etapas fetales y adultas, así como la importante regulación por la eritropoyetina y la morfología celular.








































![DVDFab Passkey Crack + License Key [Latest]](https://cdn.slidesharecdn.com/ss_thumbnails/eritropoyesis122617-250404103349-8236ca2f-250404105639-4d50982a-thumbnail.jpg?width=640&height=640&fit=bounds)


![Malwarebytes Anti-Malware Crack With Serial Key [Latest]](https://cdn.slidesharecdn.com/ss_thumbnails/eritropoyesis122617-250404103349-8236ca2f-250404105932-afea5bf0-thumbnail.jpg?width=640&height=640&fit=bounds)











































