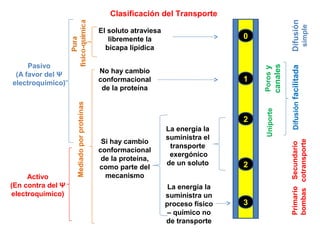
1. Clasificación del Transporte
Pasivo
(A favor del Ψ
electroquímico)
Activo
(En contra del Ψ
electroquímico)
Mediadoporproteínas
El soluto atraviesa
libremente la
bicapa lipídica
No hay cambio
conformacional
de la proteína
Si hay cambio
conformacional
de la proteína,
como parte del
mecanismo
La energía la
suministra el
transporte
exergónico
de un soluto
La energía la
suministra un
proceso físico
– químico no
de transporte
Difusión
simple
Difusiónfacilitada
Porosy
canales
Uniporte
Secundario
cotransporte
Primario
bombas
1
2
0
2
3
2.
3. Gases O2, CO2,
NO, CO,N2
polares
pequeñas
sin carga
Hidrófobas Ác. grasos
Pequeñas Esteroides
Polares glucosa
grandes sacarosa
sin carga
iones K+, Na+, Ca2+,
Mg2+. Cl-,
HCO3-, H2PO4
polares aminoácidos
cargadas glucosa--6-P
ATP, nucleótidos
H2O,urea
etanol, glicerol
Ác. Acético
ImpermeablePermeable
Permeabilidad de las membranas
4. Velocidad de difusión
Depende de:
La magnitud del gradiente de concentración.
•A mayor gradiente mejor será la difusión
Permeabilidad de la membrana.
•Membrana neuronas 20 veces más permeable al K+
que al Na+
.
Temperatura
•A mayor Tª, mayor velocidad
La superficie de difusión
•Microvellosidades incrementan el área de difusión.
5. Factores que influyen en la velocidad de difusión (j)
(nº de moléculas/unidad de tiempo)
Concentración
fuera de la cél.
Concentración
dentro de la cél.
Gradiente de
concentración
Grosor de la
membrana
Liposolu
bilidad
Tamaño
molecular
Líquido intracelular
Líquido extracelular
Composición de la
capa lipídica
Superficie de
la
membrana
J =
Área de superficie disponible x Gradiente de concentración
Resistencia de la membrana x Grosor de la membrana
Ley de Fick
6. Difusión: camino al azar (desplazamiento unidimensional)
N=nº de pasos
L=longitud del paso elemental
Desplazamiento cuadrático medio
Definimos cte de difusión
El desplazamiento cuadrático medio en un
camino aleatorio unidimensional crece
linealmente con el tiempo
En tres dimensiones
7. Estado estacionario
(J cte con tiempo)
1ª ley Fick
Estado no estacionario
(J no cte con tiempo)
2ª ley Fick
Proceso de Difusión
Macroscópicamente: difusión = f (tiempo) conocer “velocidad”⇒
Flujo de Difusión (J):
cantidad de masa (nº de átomos) M que difunden perpendicularmente a
través de un área (A) de una membrana por unidad de tiempo t
Unidades
(Kg/m2
×s; ó,
átomos/m2
×s)
14. En el equilibrio
ΔG = 0
Ψm = Ψeq
Ecuación de Nernst
R = 8,315 J·mol-1
·K-1
F = 96.480 Cb·mol-1
(J·mol-1
·V-1
)
T= 310 ºK
Walther Nernst
1864-1941
Premio Nobel 1920
CUIDADO
La ecuación de Nernst no es aplicable
a las células ya que las membranas son
permeables a mas de un ión (excepto
cél. Gliales: permeables solo a K+
)
La Ecuación de Nernst
15. La Ecuación de Nernst
in
out
X
X
X
zF
RT
E
][
][
ln=
valencia
del ion X
potencial eléctrico
del ion X
concentración extracelular
concentración intracelular
i
o
X
X
X
z
E
][
][
log
058.0
=A 20° C, Em (en voltios) es:
RT/F es constante ( 0.058 V a 20° C y 0.061 V a 38° C)
17. Difusión de iones: potencial de membrana
Membrana impermeable al Na+
, K+
y al Cl-
Em = 0 mV
Potenciómetro
18. Potencial de Equilibrio de Nernst para el Na+
Calcular el potencial de equilibrio de Nernst para Na+
:
1. Asumir que [Na+
]o es 10 veces mayor que [Na+
]i
2. [Na+
]o/[Na+
]i = 10
3. Valencia = 1
i
o
Na
Na
Na
E
][
][
log058.0 ×=
i
o
Na
Na
Na
z
E
][
][
log
058.0
=
=+×= )1(058.0NaE
=×= )10log(058.0NaE
0.058V
58 mV
CanalesdeNa+
Separación de cargas a
través de la membrana
Membrana permeable al Na+
ENa = +58 mV
19. Potencial de Equilibrio de Nernst para el K+
Calcular el potencial de equilibrio de Nernst para K+
:
1. Asumir que [K+
]i es 10 veces mayor que [K+
]o
2. [K+
]o/[K+
]i = 0,1
3. Valencia = 1
i
o
K
K
K
E
][
][
log058.0 ×=
=×= )1.0log(058.0KE
=−×= )1(058.0KE -0.058V
i
o
K
K
K
z
E
][
][
log
058.0
=
-58 mV
CanalesdeK+
Membrana permeable al K+
Separación de cargas a
través de la membrana
Ek= -58 mV
20. Potencial de Equilibrio de Nernst para el Cl-
Calcular el potencial de equilibrio de Nernst para Cl-
:
1. Asumir que [Cl-
]o es 10 veces mayor que [Cl-
]i
2. [Cl-
]o/[Cl-
]i = 10
3. Valencia= -1
i
o
Cl
Cl
Cl
z
E
][
][
log
058.0
=
i
o
Cl
Cl
Cl
E
][
][
log058.0 ×−=
=×−= )10log(058.0ClE =×−= 1058.0ClE -0.058V
-58 mV
21. Equilibrio Donnan
En EQUILIBRIO, la relación entre iones de ambos lados de la membrana es:
Si una membrana separa dos soluciones de electrolito; de las cuales una
CONTIENE UN IÓN NO DIFUSIBLE , mientras que los otros iones difunden
libremente. Resulta una distribución desigual de los iones difusibles a
ambos lados de esta membrana. La mayor cantidad de iones se hallan en el
lado que contiene la variedad no difusible.
1)Distribución asimétrica de iones (pero se mantiene la neutralidad)
[Cl-
]i < [Cl-
]e; [K+
]i > [K+
]e; [Na+
]i > [Na+
]e
Más aniones fuera : más cationes dentro
2)Diferencia de presión osmótica
∑[ión]i > ∑[ión]e ; [Osmolaridad]i > [osmolaridad]e
3) Potencial de membrana
22. Ejemplo: Equilibrio de Donnan
__Datos
__Calculado
Potencial de
Donnan
Por tanto el potencial
electroestático para
todos será igual a:
Célula (mM) Fuera (mM) [ ]o /[ ]i
Na+
210 140 0,66
K+
15 10 0,66
Cl-
100 150 0,66
X-
≈125 0
∑q (CNa + Ck) – (CCl+ X) = 0 CNa + Ck - CCl= 0 Neutralidad
∑iones 325mM > <300 mM 25
ósmosis
23. La célula NO ES UN SISTEMA EN
EQUILIBRIO.
La célula ES UN SISTEMA EN
ESTADO ESTACIONARIO
El transporte activo aleja a la célula
del equilibrio de Donnan
24. a) Electroneutralidad: Las soluciones intra y extracelulares deben ser
eléctricamente neutras. El número de aniones debe ser igual al de
cationes a ambos lados de la membrana. El leve exceso de aniones en el
lado interno de la membrana y de cationes en el lado externo, que son
los que generan el potencial de membrana de reposo, es casi
despreciable respecto de la cantidad total de iones presentes en el medio
intra y extracelular
b) La célula debe estar en balance osmótico, para que no se hinche o
se encoja (fenómenos osmóticos).
c) En ESTADO ESTACIONARIO hay un flujo neto que siempre se
mantiene constante por determinados mecanismos Homeostásicos (se
usa la ecuación de Goldman).
d)Si la célula fuera un sistema EN EQUILIBRIO no debería haber flujo
neto transmembrana (se usaría la Ec. de Nernst)
Condiciones del entorno para que se establezca el estado estacionario,
que es el Potencial de Reposo.
25. 1) La bomba
electrogénica Na+
.K+
genera el potencial de
membrana
2) Los iones tienden a
desplazarse a favor de
su gradiente
electroquímico, a través
de la membrana
polarizada
El flujo de K+ a través de los canales de K+ de reposo; desde el citosol al
exterior celular a favor de su gradiente de concentración, el que genera el
potencial de membrana negativo en el interior. Por tanto, son los canales de
K+ de reposo, que están siempre abiertos, son los que definen la magnitud
y el signo del potencial de reposo.
El potencial de membrana es, -70 mV, y el
grosor es de 3.5 nm. El gradiente de voltaje
que se genera en esta membrana resulta de
0.07 V por 3.5x10-7
cm = 200000 V/cm.
26. Ecuación de Goldman ó de Goldman-Hodgkin-Katz
• Calcula el potencial de equilibrio cuando la membrana es permeable
a más de un ion.
• Incorpora los coeficientes de permeabilidad de cada ion (especifico
de cada membrana)
⋅⋅⋅
++
++
= −++
−++
oCliNaiK
iCloNaoK
ions
ClPNaPKP
ClPNaPKP
F
RT
E
][][][
][][][
ln
26
(facilidad con la cual un ion puede cruzar una unidad de área (1 cm) de
membrana impulsado por una diferencia de concentración 1 M. Es
proporcional al número de canales iónicos abiertos y al número de iones
que cada canal puede conducir por minuto. Unidades cm/s. La
permeabilidad no se mide directamente, sino que se suele medir el número
de canales abiertos y la conductividad de cada canal.)
27. PMR de una neurona
En las neuronas hay mas ANIONES que CATIONES (en el exterior
ocurre al revés)
Hay mayor cantidad de cargas negativas en el interior.
Se crea una diferencia de potencial a lo largo de la membrana
llamado POTENCIAL DE MEMBRANA EN REPOSO.(PMR= -70 mV)
[Na+
] 10
[K+]
140
[Cl-
] 7
[P-
] 165
[Na+
] 142
[Cl-
] 103
[K+
] 5
28. PMR
• En reposo, las permeabilidades (relativas al K+
) de los iones mas
importantes son:
PNa= 0,03
PCa= 0,001
PCl= PK= 1
∆V= - 0,058 V . Log 140 + (0,03)10 + (0,001)0,001 + 103 = -0,058 V.log 275,4 =
5 + (0,03)142 + (0,001)5 + 7 14,3
∆V = -0,071 V = - 71 mV
⋅⋅⋅
++
++
= −++
−++
oCliNaiK
iCloNaoK
ions
ClPNaPKP
ClPNaPKP
F
RT
E
][][][
][][][
ln
28
INTRACELULAR
mEq/L
EXTRACELULAR
meq/L
Na+
10 142
K+
140 5
Cl-
7 103
Ca+2
0,001 5
29. 140 mM K+
4 mM K+
12 mM Na+
150 mM Na+
4 mM Cl-
120 mM Cl-
Interior Exterior
Canal de
reposo K+
Canal de
reposo Na+
Canal de
reposo Cl-
Membrana
plasmática
148 mM A-
34 mM A-
Mayoría de
canales de
K+ abiertos
Escasos
canales de
Na+ , Cl-
abiertos
Potencial de reposo
30. 0
+50
-50
+100
-100
Hiperpolarización→
ENa+ = +64,7 mV
Ereposo = -52,9 mV
Ecl- = -87,2 mV
EK+ = -91,1 mV
Efectos de las variaciones de permeabilidad iónica sobre el potencial de
membrana, usando las constantes de permeabilidad. El potencial de membrana en
reposo es de -53 mV; ENa+ , EK+ y ECl- son los potenciales calculados a partir de la
ecuación de Nernst si la membrana solo contiene canales abiertos para Na+
K+
o CI-
,
respectivamente.
PNa+ x 200
PNa+ x 0
PK+ x 200
PK+ x 1/5
PCl- x 200
PCl- x 0
32. A C
D
E
B
NPA
NPA
C189
HgCl2
Dos tripletes NPA: AQP1 se compone de dos grandes
dominios, imagen especular uno del otro. Se organiza
en seis segmentos de estructura α-hélice (1-6) que
atraviesan la membrana de lado a lado; están unidos
por cinco lazos conectores (A-E). El lazo intracelular B
y el extracelular E son portadores de un triplete
asparagina-prolina-alanina NPA. Esta duplicación del
triplete NPA es característica de todas las
acuoporinas
Se han identificado 12 acuaporinas (AQPs) se clasifican en 2 subfamilias:
Acuaporinas: canales capaces de transportar agua.
Acuagliceroporinas: canales permeables al agua y otros pequeños
solutos, como urea o glicerol. Las AQP3, AQP7, AQP9 y AQP10 pertenecen
al grupo de las acuagliceroporinas.
DE CUATRO EN CUATRO
Aunque cada acuaporina forma por
sí sola un canal, en la membrana
celular estas proteínas se
ensamblan en grupos de cuatro.
Parece que un arreglo tetramérico
confiere a la estructura una mayor
estabilidad en el entorno lipídico de
la membrana.
33. EL RELOJ DE ARENA El lazo B y E se pliegan hacia la membrana
para formar el poro o canal acuoso de la proteína. La estructura
resultante encierra una zona central estrecha que se ensancha
abriéndose hacia ambos lados de la membrana. Se denominó “reloj
de arena”, por su semejanza con éste. En el sitio más estrecho del
poro acuoso se ponen en contacto los tripletes NPA.
34. This cartoon of bovine aquaporin 1
shows that the pore (brown; filled with
water molecules shown in red and
white) narrows at His180 to a diameter
of 2.8 Å (about the size of a water
molecule), limiting passage of
molecules larger than H2O. The positive
charge of Arg195 repels cations,
including H3O+, preventing their
passage through the pore. The two
short helices shown in green are
oriented with their positively charged
dipoles pointed at the pore in such a
way as to force a water molecule to
reorient as it passes through; this
breaks up hydrogen-bonded chains of
water molecules, preventing proton
passage by "proton hopping"
Size
restriction
Electrostatic
repulsion
Waterdipole
reorientation
36. Son proteínas transmembrana que contienen un canal o poro de naturaleza
polar; que permite el paso de iones, a favor de su gradiente electroquímico
cuando está abierto
Están constituidos por subunidades homólogas o idénticas, o por dominios
homólogos de una única cadena polipeptídica.
Presentan diversas conformaciones con diferentes conductancias iónicas:
un estado cerrado y uno abierto, y algunos, estados inactivos controlados
por temporizadores internos.
Los canales iónicos
Los estados no conductores son los
más estables termodinámicamente
37. Presentan selectividad. Sobre lo cual influye las cargas en la pared del
poro así como el diámetro de éste.
Mecanismo de apertura y cierre (estímulos diversos)
Clasificación según estímulo que incrementa la probabilidad de apertura
(Po):
Por voltaje,
Por estiramiento (mecanosensibles),
Por calor
Por depósito (Ca2+)
Por ligandos:
1)mensajeros extracelulares (receptores ionotrópicos);
2) 2º mensajeros, (receptores metabolotrópicos); (proteínas Gβγ,
AMPc, GMPc, IP3 ó Ca2+
La fosforilación puede modificar su actividad
38. Canal de K+
En reposo
Siempre
abierto
Canal regulado
por voltaje
Apertura (transitoria)
en respuesta a una
variación del
Potencial de
membrana
Canal regulado por ligando
Apertura (cierre) en
respuesta a un
Neurotransmisor
extracelular
específico
Apertura (cierre) en
respuesta a una
molécula
intracelular
específica
Receptor
Inotrópico
Receptor
Metabolotrópico
GRADIENTES
Interior
Exterior
39. Tipo funcional Ligando Canal iónico
Excitatorios
Acetilcolina (nicotinico) Na+
/K+
Glutamato (NMDA) Na+
/K+
y Ca2+
Glutamato (no NMDA) Na+
/K+
Serotonina (5HT3) Na+
/K+
Inhibitorios
Ac. γ-aminobutirico, GABA (clase A) Cl-
Glicina Cl-
Receptores ionotrópicos.
41. Los canales iónicos
Operados por voltaje son muy selectivos: p.ej. los de K+ son 100 a
1000 veces más permebles al K+ que al Na+. Pero los de Na+ 11 a 20
veces más al Na+ que al K+. Los operados por ligando seleccionan
pobremente.
Pueden entrar en estados desensibilizados, refractarios o inactivos
(importante en codificación temporal y protección ante estimulación
excesiva):
En aquellos activados por ligando: ocurre después de prolongada
exposición al ligando.
En aquellos activados por V: para los de Na+, Ca2+ y K+: por el
mismo cambio de V. Los de Ca2+: también por influjo del mismo
calcio.
42. 6 subunidades
4 subunidades, algunos
con otra β
4 subunidades, algunos con otra β
(7DT) (facilitan la salida de K+ →
hiperpolarización, ↓la frecuencia
de disparo
4 subunidades (actúan en las
hiperpolarizaciones, permiten la
entrada de K+ → Despolarización
Clasificación de los canales iónicos (1)
24 familias (256 genes)
4 subunidades, algunos con otra β
(salida de K+ durante el potencial
de acción → hiperpolarización
43. Clasificación de los canales iónicos (2)
4 subunidades
4 subunidades
4 subunidades
2 subunidades
(potencial de reposo)
44. Clasificación de los canales iónicos (3)
2 o 3 subunidades (α1, β1 y β2)
5 subunidades (α1, β, α2-δ y γ)
47. Las familias de los canales de potasio:
(A)dependientes de voltaje.
(B) los canales de potasio con dos poros.
(C) los dependientes de Calcio.
(D)los rectificadores internos (D).
48. Canal de K+
bacteriano Streptomyces lividans.
La estructura del canal de K+
: está formado por cuatro subunidades idénticas que abarcan la
membrana y forman un cono dentro de otro cono en torno al canal iónico, con el extremo el
doble de ancho mirando hacia el espacio extracelular. Cada subunidad tiene dos hélices α
transmembrana y una tercera hélice más corta que contribuye a la región del poro. El cono
externo está formado por una hélice transmembrana de cada subunidad. El cono interior,
formado por las cuatro hélices transmembrana restantes, rodea el canal iónico y contiene el
filtro de selectividad de ión. La longitud del canal es de 34 Å, el diámetro del poro inicial es de
10 Å, pero se va estrechando hasta llegar a los 3 Å, en una región de 12 Å.
49. Oxígenos carbonílicos de la
cadena principal forman una
jaula a la que se adapta el K+ de
forma precisa, reemplazando el
agua de la capa de solvatación
Sitios alternantes de K+
(azul ó verdes) ocupados
El dipolo de la hélice, y
otras cargas negativas ↑
La [k+] y lo estabilizan
El amplio vestibulo
lleno de agua permite
la solvatación del K+K+ con la capa de
solvatación
El reloj de arena
34Å
10 Å
3 Å
12Å.
50. Especificidad = Selectividad
En el filtro de selectividad existen cuatro
sitios potenciales de fijación de K+
, cada
uno de ellos formado por una "jaula" de
oxígeno que proporciona sitios de unión
para los iones K+
.
51. ΔG = -
ΔG = +
Debido a que el Na+
es demasiado
pequeño para interaccionar
favorablemente con el filtro de
selectividad la energía libre de la
desolvatación no puede
compensarse y el Na+
no pasa a
través del conducto. El
movimiento del K+ en el canal de
K+
se realiza a favor de gradiente.
52. Avance dentro del canal
Por alternancia de las conformaciones 4-2 y 3-1
Equilibrio
EquilibrioRepulsión
Repulsión
Coformación
4-2
Coformación
4-2
Coformación
3-1
54. Canal de K+ de compuerta regulada por de voltaje (shaker)
Subunidad β
Subunidad α
Estructura del complejo formado por la
subunidad α Kv1.2 y la subunidad auxiliar β2
DominioT
Subunidad α
Subunidad β
DominioT
56. Mecanismo de cierre y apertura del canal
(A) Modelo convencional en el que el movimiento de cargas se realiza a través del
núcleo proteico del canal mediante un movimiento de traslación y/o rotación del S4.
(B) Modelo en el que el sensor de voltaje se encuentra sumergido en los lípidos
de la membrana, produciendo el desplazamiento de cargas a través del campo
eléctrico. [Adaptadas de Jiang y cols., 2003b]
la secuencia de cada S4 contiene cuatro o más residuos cargados positivamente
(argininas y lisinas) que se recolocan en la membrana cuando el Em varía produciendo
un movimiento de cargas a través del campo eléctrico transmembrana que se puede
medir experimentalmente (≈12-13 eo)
(A)
(B)
57. Mecanismo de inactivación
La inactivación tipo-N se conoce como “modelo de la bola y la cadena”. En este modelo, la
bola (una secuencia de aproximadamente 20 aminoácidos del extremo N-terminal) ocluye el
poro intracelular del canal e impide el paso de los iones a su través. Los aminoácidos que
unen esta bola al S1 forman la denominada cadena. La inactivación tipo-N es un proceso muy
rápido (≈1-10 ms), por lo que también se conoce como “inactivación rápida” y se produce en
canales de la familia Shaker (Kv1.1 y Kv1.4), Shal (Kv4.2 y Kv4.3) y Shaw (Kv3). Además,
existen diferentes subunidades auxiliares (como las subunidades Kvβ) que pueden
interaccionar con los canales, ocluyendo el poro y dando lugar a una inactivación tipo-N de los
mismos.
58. Diferentes conformaciones en las que se puede encontrar el canal
- - - - - - - - -
Polarizada
+ + + + + + + +
Despolarizada
+ + + - - - - - - - -
+ + +
+ + +
Entrada del poro - - -
59. Canal de Na+
regulado por voltaje
El componente esencial de un canal de Na+
es un polipéptido largo y sencillo (1840
residuos aminoácidos) organizado en cuatro dominios agrupados alrededor de un
canal central, proporcionando un paso para el Na+
a través de la membrana. Este paso
es específico para Na+
gracias a una "región de poro" compuesta por segmentos entre
las hélices transmembrana 5 y 6 de cada dominio, que se pliegan dentro del canal
P
P
P
P
β1β2
Extracllular
space
Cytosol
Site of cAMP-dependent
Protein phosphorilation
60. (A) Estado
de reposo
con canal
de Na+ activo
cerrado
Despolarización
Inicial < 0,1 ms
(b) Desplazamiento
De la hélice α
y apertura del canal
(c) Canal inactivo
(periodo
refractario)
0,5-1,0 ms
Membrana despolarizada
Poro
selectivo
De ion
Na+
Hélice α
detectora del
voltaje
Portal
Vestibulo
interno
Segmento inactivador
del canal
Vestibulo
externo
1) Repolarización de la membrana
2) Desplazamiento del segmento inactivador del canal
3) Cierre de la compuerta
Lento (varios ms)
Exterior
Citosol
61. Voltage-gated Na+ channels of neurons. Sodium channels of different tissues and organisms
have a variety of subunits, but only the principal subunit (α) is essential. (b) The four domains
are wrapped about a central transmembrane channel lined with polar amino acid residues. The
four pore regions (red) come together near the extracellular surface to form the selectivity
filter, which is conserved in all Na+ channels. The filter gives the channel its ability to
discriminate between Na+ and other ions of similar size. The inactivation gate (green) closes
(dotted lines) soon after the activation gate opens.
62.
63. 1 A stimulus to this neuron (not shown) causes an
action potential to move along the axon (white
arrow), away from the cell body. The opening of a
voltage-gated Na+ channel allows Na+ entry, and
the resulting local depolarization causes the
adjacent Na+ channel to open, and so on. The
directionality of movement of the action potential is
ensured by the brief refractory period that follows
the opening of each voltage-gated Na+ channel. 2
When the wave of depolarization reaches the axon
tip, voltage-gated Ca2+ channels open, allowing
Ca2+ entry. 3 The resulting increase in internal
[Ca2+] triggers exocytic release of the
neurotransmitter acetylcholine into the synaptic
cleft. 4 Acetylcholine binds to a receptor on the
postsynaptic neuron (or myocyte), causing its
ligand-gated ion channel to open. 5 Extracellular
Na+ and Ca2+ enter through this channel,
depolarizing the postsynaptic cell. The electrical
signal has thus passed to the cell body of the
postsynaptic neuron (or myocyte) and will move
along its axon to a third neuron (or a myocyte) by
this same sequence of events.
64. Activación secuencial de canales iónicos regulados en una unión neuromuscular. La llegada de
un potencial de acción a la terminal de una neurona motora presináptica induce la apertura de
los canales de Ca2+
regulados por voltaje 1 y la liberación posterior de acetilcolina que induce la
apertura de los receptores nicotínicos regulados por ligando de la membrana muscular 2. El
consecuente ingreso de Na+
produce una despolarización localizada de la membrana que induce
la apertura de canales de Na+
regulados por voltaje y la generación de un potencial de acción 3.
Cuando la despolarización se difunda hasta alcanzar los túbulos T, desencadena la apertura de
los canales de liberación de Ca2+
regulados por voltaje y la liberación de Ca2+
desde el retículo
sarcoplásmico hacia el interior del citosol 4 el incremento de Ca2+
citosólico produce la
contracción muscular.
65. The acetylcholine receptor ion channel. (a) Each of the five homologous subunits (α2βγδ) has
four transmembrane helices, M1 to M4. The M2 helices are amphipathic; the others have mainly
hydrophobic residues. (b) The five subunits are arranged around a central transmembrane
channel, which is lined with the polar sides of the M2 helices. At the top and bottom of the
channel are rings of negatively charged amino acid residues. (c) A model of the acetylcholine
receptor, based on electron microscopy and x-ray structure determination of a related protein
(the acetylcholine-binding protein from a mollusk).
(a) Cada subunidad se pliega
en cuatro hélices α
transmembrana: M1, M2, M3 y
M4; con secuencias
extramembrana dentro y
fuera
(b) Las hélices M1, M2 y
M3 son hidrofóbicas; Las
M2 son anfipáticas y
rodean el canal
(c) Sitios de
unión de la
acetilcolina
Receptor de acetílcolina : composición y estructura
Exterior
Interior
66. Características del canal
La mayor parte de la proteína sobresale de la superficie externa de la membrana plasmática.
La hélice M2 (rojo) de cada subunidad es parte del revestimiento del canal iónico. Las
cadenas laterales de aspartato y glutamato, en ambos extremos de cada hélice M2, forman
dos anillos de carga negativa que contribuyen a excluir aniones del canal y atraer cationes.
La compuerta que se abre por la fijación de acetilcolina se encuentra dentro del poro
67. Apertura y cierre del canal
Cadenas laterales
voluminosas e hidrofóbicas
de Leu en las hélices M2,
cierran el canal
Las hélices M2 tienen ahora
residuos polares más pequeños
recubriendo el canal abierto
La unión de 2 moléculas
de acetilcolina produce
un giro en las hélices M2
72. Estructura de transportadores Glut
PM = 45000 Da
12 segmentos transmembrana
8 hélice anfipáticas (polares sin carga; forman puentes de hidrógeno)
73. Glucose transporter 1 (GLUT1) structure. A and B: GLUT1 structure as modeled in Ref. 10 and drawn
using VMD (version 1.8.5). The 12 transmembrane helices are shown in 2 forms, as a surface
representation (left) or as a cartoon representation (right). Extracellular and cytoplasmic structures are
omitted. The cartoon representations include the transmembrane helix numbers. A: transmembrane α-
helices (TMs) are shown parallel to the membrane. B: the TMs are viewed along the membrane normal
from the cytoplasmic side.
74. Model for GLUT1-mediated sugar
transport. A: schematic
representation of the catalytic
center of the transporter.
Extracellular sugar (Go) and
intracellular sugar (Gi) react with
exo- and endofacial sites,
respectively, to form G2 and G1,
respectively. Sugar dissociates
from these sites into the intersite
cavity to form Gc. When G2 and G1
are occupied, dissociation to Gc
and reassociation are accelerated
B: The GLUT1 model (TMs 2 and
11 removed for clarity) showing Gi
complexed to the G1 site, a
putative G2 site, the small intersite
cavity,
La tetramerización parece ser la asociación más probable
75.
76. Cinética del transporte de glucosa a los eritrocitos
Concentración extracelular
de glucosa [S]out (mM)
Velocidadinicialdeentradade
glucosaVo(μM/min)
Monosacárid
o
Especificidad
(Kt) (mM)
D- Glucosa 1,5
D- Manosa 20
D-
Galactosa
30
L- Glucosa >3000
Características del transporte pasivo
Velocidad de difusión elevada
Saturable
Especificidad
77. Otros transportadores de glucosa
En el genoma humano se encuentran codificados doce transportadores de
glucosa (se diferencian en: propiedades cinéticas, distribución tisular y función
Transportador Tejidos donde se expresa Función
GLUT1
(Kt = 1,5 mM)
Ubicuo Captación basal de glucosa
GLUT2
(Kt = 66 mM)
Hígado, intestino y páncreas Eliminación de la
hiperglucemia (hígado),
control de liberación de
insulina (páncreas)
GLUT3 Cerebro Captación basal de glucosa
GLUT4
(Kt = 5 mM)
Músculo, Tej. adiposo y
corazón
Dependiente de insulina
GLUT5 Intestino, riñón Principalmente transporte de
fructosa
82. Two types of active transport. (a) In primary active transport, the energy released by
ATP hydrolysis drives solute movement against an electrochemical gradient. (b) In
secondary active transport, a gradient of ion X (often Na+) has been established by
primary active transport. Movement of X down its electrochemical gradient now
provides the energy to drive cotransport of a second solute (S) against its
electrochemical gradient.
Transporte activo
83.
84. Transporte activo secundario: Cotransporte sodio-glucosa en el intestino
(simporte)
Glucose transport in intestinal epithelial cells. Glucose is cotransported with Na+ across the apical
plasma membrane into the epithelial cell. It moves through the cell to the basal surface, where it
passes into the blood via GLUT2, a passive glucose uniporter. The Na+K+ ATPase continues to
pump Na+ outward to maintain the Na+ gradient that drives glucose uptake.
86. The primary transport of H+
out of the cell, driven by the oxidation of a variety of fuels,
establishes both a proton gradient and an electrical potential (inside negative) across the
membrane. Secondary active transport of lactose into the cell involves symport of H+
and
lactose by the lactose transporter. The uptake of lactose against its concentration gradient is
entirely dependent on this inflow of protons driven by the electrochemical gradient.
Lactose uptake in E. coli.
87. Lactose uptake in E. coli.
When the energy-yielding oxidation reactions of metabolism are blocked by cyanide (CN–
), the
lactose transporter allows equilibration of lactose across the membrane via passive transport.
Mutations that affect Glu325
or Arg302
have the same effect as cyanide. The dashed line
represents the concentration of lactose in the surrounding medium.
88. Cuatro clases de ATPasas transportadoras
Organismo/tejido Membrana Función
ATPasas tipo P
Na+
-K+
Animal Plasmática Mantiene baja la [Na+
] y
alta la [K+
] dentro de la
célula. Crea un potencial
eléctrico transmembrana
H+
-K+
Células (parietales)
secretoras de ácido
de mamífero
Plasmática Acidifica el contenido del
estómago
H+
Hongos Plasmática Crean un gradiente de
[H+
];para transporte
activo secundario de
solutos, hacia
dentro de la célula
H+
Plantas superiores Plasmática
Ca2+
Animal Plasmática Mantiene baja la [Ca2+
] en
el citosol
Ca2+
Miocitos Retículo
sarcoplasmico
Secuestra Ca2+
Intracelular, manteniendo
baja la [Ca2+
] intracelular
Cd2+
,Hg2+
, Cu2+
Bacterias Plasmática Bombean metales
pesados fuera de la célula
89. Composición:
Tetrámero de α2β2 (+ γ en riñón regulatoria)
Tiene 2 conformaciones E1 y E2
β: Muy glicosilada, plegamiento e inser. de las subunidades α, Pm = 55 Kda
α : No glicosilada, Pm =110 Kda,10 hélices transmembrana, Asp 269
Fosforilación, Tiene las actividades ATPasa y de translocación
Inhibida por ouabaína (entre H2 y H3) (y vanadato, como toda la familia P)
Na+ - K+ ATPasa
NBD
P
Subunidad α
90. Structure of
ouabain, a
cardiotonic steroid,
a potent inhibitor of
the Na+/K+
exchanging ATPase.
Proposed sequence of reactions and
intermediates in hydrolysis of ATP by
the Na+/K+ exchanging ATPase.
91. Mecanismo
Conformación E1
>Afinidad por Na+
< Afinidad por K+
Conformación E2
<Afinidad por Na+
> Afinidad por K+
Transiciones
Fosforilación E1 → E2
Defosforilación E2 → E1
92. Cuatro clases de ATPasas transportadoras
Organismo/tejido Membrana Función
ATPasas tipo V
H+
Animal Vesículas lisosómicas,
endosómicas y
secretoras Crean un compartimento
ácido, activando proteasas y
otros enzimas hidrolíticosH+
Plantas superiores Vacuolar
H+
Hongos Vacuolar
ATPasas tipo F
H+
Eucariotas Mitocondrial interna
Catalizan la formación de
ATP, a partir de ADP y Pi
H+
Plantas superiores Tilacoides
H+
Procariotas Plasmática
93.
94. Cuatro clases de ATPasas transportadoras
Organismo/tejido Membrana Función
Proteínas Transportadoras ABC (Multidrug Transporter)
Células tumorales Plasmática Eliminan muchos productos
animales hidrofóbicos naturales y
fármacos sintéticos del citosol; entre
ellos vimblastina, doxorubicina,
actinomicina D, mitomicina, taxol,
colchicina y puromicina (bombas)
Células epiteliales Plasmática Expulsan al exterior sal y agua
(canales)
Macrófagos
(alveolos
pulmonares)
Vesículas
lisosómicas
Ayudan a acidificar los lisosomas por
compensación de carga
95. Están en la membrana plasmática y membranas intracelulares
Estructura modular un dominio con seis segmentos transmembrana; y
un dominio citosólico de unión a ATP (NBD).
La mayoría poseen una repetición en tanden de dos dominios
transmembrana y dos dominios de unión a ATP (1 Sólo polipéptido) ó dos
polipéptidos, cada uno con un NBD y 6 ó 10 hélices transmembranas
Funcionan como bombas o como canales en contra de gradiente
Calalizan la entrada o salida de:
Lípidos [fosfolípidos, acil-CoA (cadena larga), sales biliares y
colesterol], péptidos, moléculas orgánicas tóxicas y agentes
quimioterápicos. En bacterias (de todo).
Los transportadores ABC utilizan ATP para impulsar el transporte activo
de una amplia gama de sustratos
96. Familia de las P-glicoproteínas
Multidrug resistance family (Mdr) of genes (extrusión de
xenobióticos en células tumorales
Bombeo hacia los canalículos de fosfatidilcolina, colesterol y
sales biliares desde los hepatocitos.
Etc.
Multidrug resistance family (Mdr)
98. Es un canal regulador de conductancia y un canal de Cl-
Posee 5 dominios en un polipéptido: 2 dominios con seis hélices
transmembrana, 2 dominios de unión a ATP (NBD) y un dominio regulador
Interviene en los movimientos de sal y agua en la membrana apical
epitelial
CFTR cystic fibrosis transmembrane conductance regulator
P
La mutación ocasionada por la
delección de Phe 508 en NBD1
es la causante de la mayoría de
casos de fibrosis quística
99. Mecanismo ?
Three states of the cystic fibrosis transmembrane conductance regulator, CFTR. The protein has two segments,
each with six transmembrane helices, and three functionally significant domains extend from the cytoplasmic
surface: NBD1 and NBD2 (green) are nucleotide-binding domains that bind ATP, and a regulatory domain (blue) is
the site of phosphorylation by cAMP-dependent protein kinase. When this R domain is phosphorylated but no
ATP is bound to the NBDs (left), the channel is closed. The binding of ATP opens the channel (middle) until the
bound ATP is hydrolyzed. When the regulatory domain is unphosphorylated (right), it binds the NBD domains and
prevents ATP binding and channel opening. The most commonly occurring mutation leading to CF is the deletion
of Phe508 in the NBD1 domain (left). CFTR is a typical ABC transporter in all but two respects: most ABC
transporters lack the regulatory domain, and CFTR acts as an ion channel (for Cl–), not as a typical transporter.
Canal cerrado
Dominio R fosforilado
no hay ATP unido a los
NBD
Canal abierto
Dominio R fosforilado
ATP unido a los NBD
Canal cerrado
Dominio R sin fosforilar
no hay ATP unido a los
NBD
100. Superficie epitelial
de las vías aéreas
Glándula
submucosa
Normal CF
Ejemplo: pulmón (en el resto de conductos el efecto es similar)
101. PROCESOS QUIMIOSMÓTICOS
Cuando una membrana delimita un compartimento cerrado, y se establece por algún
mecanismo una diferencia de concentración de un soluto –o de potencial, si es el
caso- a ambos lados de la membrana, el transporte del soluto a favor de su potencial
es exergónico, esto es, libera energía libre. Esta es una situación “de alta energía de
transporte” análoga a la situación “de alta energía de hidrólisis” que tienen los
enlaces pirofosfato que se encuentran en el ATP. En ambos casos cuando el sistema
(gradiente de soluto; ATP) cambia de estado (el soluto se transporta; el ATP se
hidroliza) se libera energía que, de existir un mecanismo de acoplamiento adecuado,
se puede aprovechar. Esta similitud está en la base del modelo quimiosmótico de
Peter Mitchell, que fue quien por primera vez desarrolló la idea de la posible
transducción de energía entre gradientes de soluto –el componente "osmótico"- y
reacciones químicas.
De hecho, durante mucho tiempo se consideró al ATP la “moneda energética de la
célula”, que transfería energía libre de unas reacciones a otras. Pues bien, las células
(todas) tienen dos tipos de “moneda energética”, dos tipos de mecanismos de
almacenamiento inmediato y de transducción de energía: el sistema ATP-ADP y los
gradientes de solutos. Podemos extender esta metáfora, diciendo como V. Skulachev
que las células pueden usar “efectivo” –el ATP- y “cheques”: los gradientes de
solutos. Y ambos tipos de "dinero" son intercambiables entre sí.
102. 1: Anhidrasa carbónica
2: Antiporte electroneutro cloruro-bicarbonato
3: H+K+ ATPasa electroneutra.
4: Sinporte electroneutro cloruro-potasio
En rojo transporte endergónico; en azul, transporte exergónico
Acidificación del jugo gástrico
El ión Cl-
proviene de la sangre;
el H+
procede del agua; ambos
se deben exportar al jugo
gástrico en contra de sus
potenciales electroquímicos.
103. Circuitos quimiosmóticos en una célula animal
En una célula animal típica, la mayor parte del
ATP se sintetiza a partir de ADP y Pi mediante
la fosforilación oxidativa mitocondrial. En este
proceso los tres centros redox de la cadena
respiratoria mitocondrial bombean
hidrogeniones desde la matriz mitocondrial al
espacio intermembrana, desarrollando una
fuerza protonmotriz del orden de - 180 mV. Esa
fuerza protonmotriz permite la síntesis de ATP
mediante la ATPsintetasa F. El ATP se puede
emplear, entre otras muchas reacciones, para
establecer los gradientes de Na+
y K+
a través
de la membrana plasmática de la célula
(mediante la bomba de Na+
K+
del tipo P), y
para acidificar los lisosomas mediante una
ATPasa V. El gradiente de Na+
se emplea como
fuente de energía para el transporte de solutos
a través de la membrana plasmática mediante
diferentes procesos de cotransporte.
Obviamente esto es un modelo muy
simplificado: hay otras ATPasas de tipo P y de
tipo ABC en los diferentes sistemas de
membrana
104. Circuitos quimiosmóticos en una célula vegetal
En una célula vegetal NO fontosintética la
situación es similar a la de la célula
animal (el ATP se obtiene mediante la
fosforilación oxidativa), con la diferencia
de que el soluto que se utiliza para el
cotransporte a través de la membrana
plasmática es el ión H+
, en vez del Na+
. El
gradiente de H+
en la membrana
plasmática se crea mediante una ATPasa
de tipo P. En hongos y levaduras la
situación es similar, excepto cuando
están en anaerobiosis. En ese caso, el
ATP se produce exclusivamente por la
fosforilación a nivel de sustrato que
ocurre en la fermentación. En una célula
fotosintética existe un tercer
compartimento, el cloroplasto, que
sintetiza ATP. En este caso se crea un
gradiente de pH a través de la membrana
del tilacoide, entre el estroma del
cloroplasto y el lumen del tilacoide. El
ATP se puede emplear en los procesos
biosíntéticos del cloroplasto, o ser
exportado al citosol.