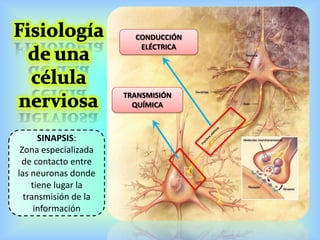
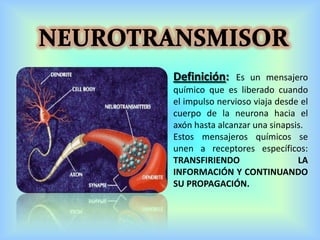
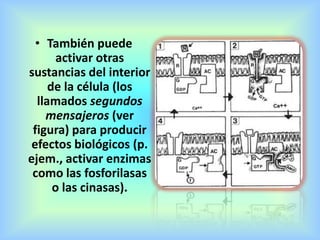
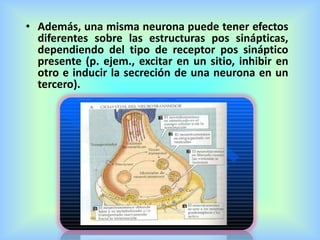
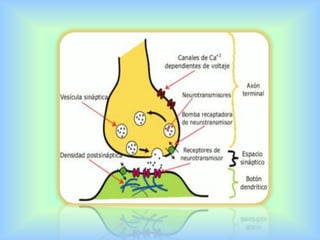
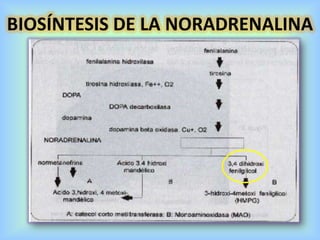
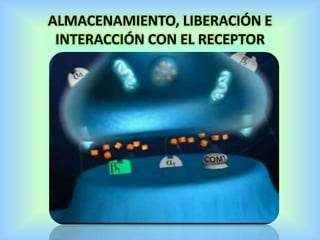
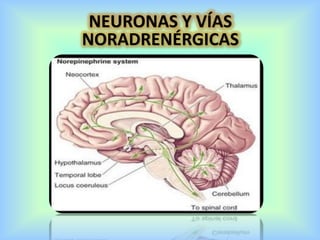
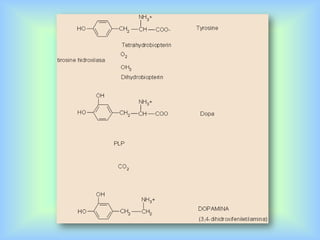
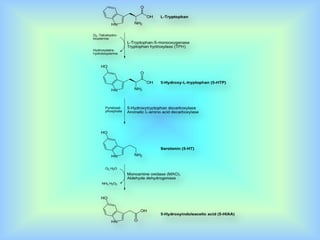
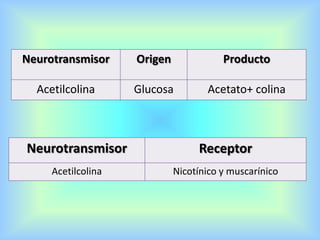
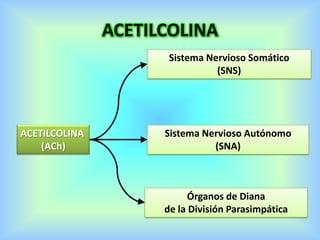
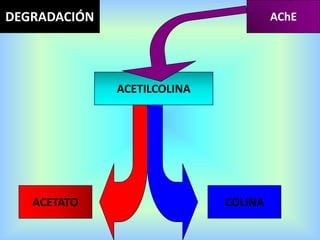
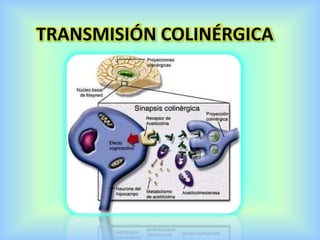
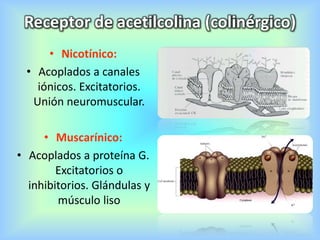
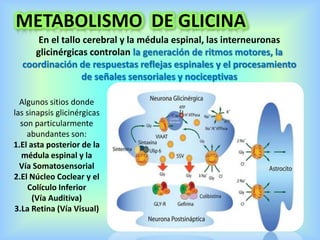
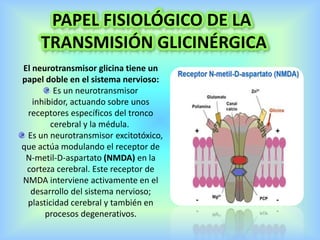
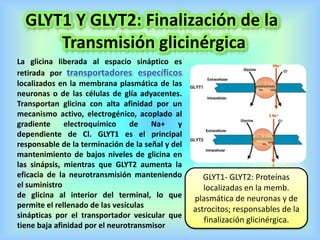
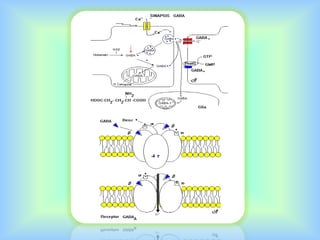
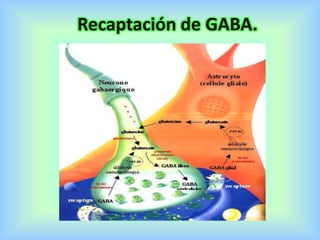
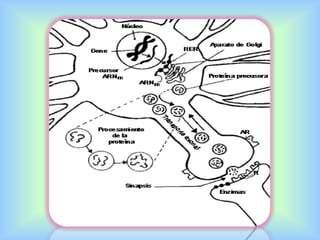
El documento describe varios aspectos de la transmisión química en las sinapsis neuronales. Explica que los neurotransmisores son sustancias químicas liberadas por la neurona presináptica que se unen a receptores en la neurona postsináptica, transmitiendo la señal nerviosa. Los neurotransmisores más comunes son la acetilcolina, el glutamato, la noradrenalina, la dopamina y la serotonina. Además, detalla los mecanismos de liberación, recaptación y degradación de los neurotransmis





















































































![[CapíTulo 6] Neurotransmisores Y Sus Receptores](https://cdn.slidesharecdn.com/ss_thumbnails/captulo6neurotransmisoresysusreceptores-090810165319-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)
















![Olga neuro terminada[1]](https://cdn.slidesharecdn.com/ss_thumbnails/olganeuroterminada1-101029135956-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)



![neuro [Autoguardado].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/neuroautoguardado-220822174614-897ff743-thumbnail.jpg?width=640&height=640&fit=bounds)
































