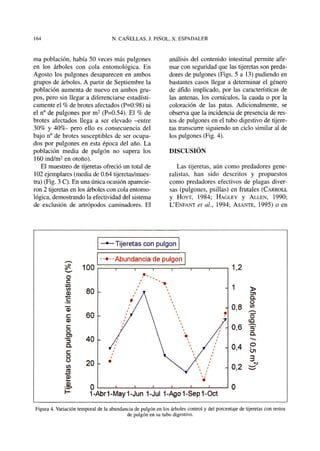
Este documento presenta los resultados de un estudio sobre la posible función de las tijeretas (Forficula auricularia) como depredadores de pulgones en un cultivo ecológico de mandarinos en España. Los resultados muestran que la presencia de tijeretas redujo significativamente las poblaciones de pulgones en comparación con los árboles donde se excluyeron las tijeretas. Además, se encontraron restos de pulgones en el tracto digestivo de las tijeretas, lo que demuestra que efectivamente los consumen