Descargado 20 veces


















































![Desarrollo y validación de un sistema de clasificación para los ríos de Cantabria
Esta técnica de clasificación está basada en la construcción de árboles de
decisión aleatorios que permiten considerar una gran cantidad de variables de
partida. Una diferencia importante entre RF y los otros métodos utilizados para la
clasificación (LDA y GDM) radica en que RF genera clasificaciones no anidadas. Por
lo tanto, en las clasificaciones dadas por RF las unidades (i.e. subcuencas) que
aparecen agrupadas en una misma clase para un determinado nivel de corte, puede
que no pertenezcan a la misma clase en un nivel de corte superior. Esto se produce
porque cada nivel de corte propuesto por RF está basado en modelos
independientes, por lo que cada nivel viene definido por un modelo predictivo
diferente, generando, finalmente, un “bosque aleatorio” frente a la clasificación en
árbol de los Classification Trees.
Además, el método multivariante RF proporciona medidas de la importancia
relativa que las diferentes variables predictoras tienen en la clasificación, entre ellas
las medidas basadas en el índice Gini (mean decreases in Gini index; MDG) o en la
precisión de las estimaciones (mean decreases in accuracy; MDA).
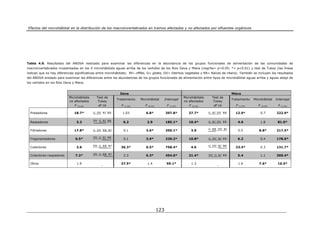
Validación de las clasificaciones
Una vez aplicados los 3 métodos de clasificación indicados se analizó la
capacidad de discriminación de las diferentes clases propuestas por los modelos.
Esta valoración se realizó mediante la aplicación de 3 análisis estadísticos: (1)
Análisis de Similaridad (ANOSIM; Clarke, 1993), (2) Análisis de Similaridad Media
(MeanSim; Van Sickle, 1997) y (3) Análisis de Homogeneidad (Homog; Bedward et
al., 1992).
1. El estadístico R del ANOSIM está basado en la diferencia de los
rangos medios entre grupos rB y dentro de los grupos rW. Así, R se define como:
R = 4(rB-rW)/ [n(n-1)]
Donde n es el número total de puntos de estudio. El valor de R varía entre
-1 y 1, donde el valor 0 indica una agrupación completamente aleatoria y los
valores positivos de R indican que las diferencias entre grupos son mayores que
dentro de los mismos.
2. El MeanSim es similar al ANOSIM, aunque utiliza el estadístico
denominado Classification Strength (CS) para medir las diferencias entre grupos.
CS se calcula como:
CS = 2Sb/ (Sw1+Sw2)
Donde Sb = similaridad media entre grupos, Sw1 = similaridad media dentro
del grupo 1 y Sw2 = similaridad media dentro del grupo 2. Si el valor de CS es >0
43](https://image.slidesharecdn.com/tesismac-121105053812-phpapp02/85/Tesis-mac-51-320.jpg)






































































































































Este documento presenta la tesis doctoral de Mario Álvarez Cabria titulada "Estudio de la variabilidad espacio-temporal de las comunidades de macroinvertebrados bentónicos en los ecosistemas fluviales de Cantabria. Repercusiones para la aplicación de la Directiva Marco del Agua". La tesis fue dirigida por José Barquín Ortiz y José A. Juanes de la Peña en la Universidad de Cantabria. El documento incluye agradecimientos y el índice de la tesis, la cual está compuesta por seis capítu



















































![Macroinvertebrados Bentonicos Como Bioindicadores De Contami[1]](https://cdn.slidesharecdn.com/ss_thumbnails/macroinvertebradosbentonicoscomobioindicadoresdecontami1-100324214238-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)
![Macroinvertebrados Bentonicos Como Bioindicadores De Contami[1]](https://cdn.slidesharecdn.com/ss_thumbnails/macroinvertebradosbentonicoscomobioindicadoresdecontami1-100326011336-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)































