Descargar para leer sin conexión
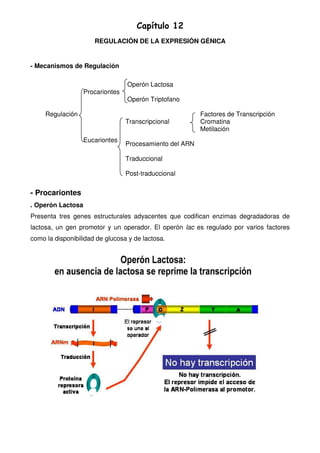









El capítulo describe los mecanismos de regulación de la expresión génica en procariotas y eucariotas. En procariotas, se explican los operones lactosa y triptófano y cómo regulan la expresión de genes en respuesta a sustratos. En eucariotas, la regulación ocurre a través de factores de transcripción, la estructura de la cromatina, la metilación del ADN, el procesamiento del ARN y los controles a nivel traduccional y post-traduccional como las chaperonas y la ubiquitinización.














































