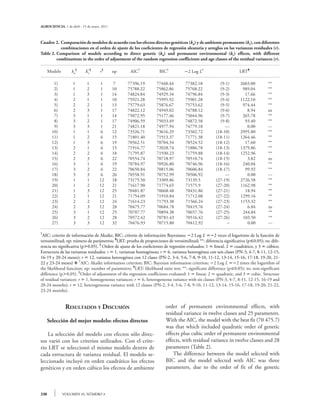
Este estudio ajusta un modelo de regresión aleatoria basado en polinomios de Legendre para evaluar el crecimiento en bovinos Tropicarne, utilizando datos de 12,890 pesadas mensuales de 1,787 becerros hasta los 24 meses. Se compararon 27 modelos para estimar efectos genéticos y de ambiente, encontrando que el modelo con mejor ajuste incluye efectos cuadráticos de genética y cúbicos de ambiente con varianza residual heterogénea. La heredabilidad directa media fue de 0.079 y la materna de 0.084, aunque el modelo subestimó los parámetros genéticos en edades más jóvenes.




![AGROCIENCIA, 1 de abril - 15 de mayo, 2011
k=4 ka−1 kc−1 1) and the covariable age of the cow, with linear and quadratic
yij = Fij + ∑ βr Φr ( aij ) + ∑ αir Φir ( aij ) + ∑ ρir Φir ( aij ) effects; aij is the jth standardized age (1 a 1) at which the ith
r=2 r=2
km−1 kq−1 animal was weighed; ir is the rth Lengendre polynomial; k is the
+ ∑ γir Φir ( aij ) + ∑ δir Φir ( aij ) + εij order of the evaluated polynomials (2 linear, 3 quadratic,
r=2 r=2 and 4 cubic), in which ka direct genetic effects, kc
animal permanent environmental effects, km maternal genetic
donde, yij es el j-ésimo peso del i-ésimo animal; Fij es el con- effects, and kq permanent maternal environmental effects; r
junto de efectos fijos en el j-ésimo registro del i-ésimo animal, is the coefficient of regression of cubic order for modeling the
que considera los grupos contemporáneos (definidos por la population mean; ir and ir are the coefficients of regression
combinación de año-época de pesada, sexo y edad del animal for modeling the direct random, genetic and permanent
en meses; el número de grupos contemporáneos por edad se environmental effects, of the rth polynomial for the ith animal;
presenta en el Cuadro 1) y la covariable edad de la vaca, con ir and ir are the coefficients of regression for modeling the
efectos lineal y cuadrático; aij es la j-ésima edad estandarizada maternal random, genetic, and permanent environmental effects
(1 a 1) a la que se pesó el i-ésimo animal, ir es el r-ésimo of the rth polynomial for the ith animal; and ij is the residual
polinomio de Legendre; k es el orden de los polinomios eva- effect. The cubic order for modeling the population mean ()
luados (2 lineal, 3 cuadrático y 4 cúbico), en el que was defined in preliminary analyses considering only fixed effects
ka efectos genéticos directos, kc efectos de ambiente per- of year and time of birth and sex of the animal as classes, plus
manente del animal, km efectos genéticos maternos y kq the covariable animal age in linear, quadratic and cubic function.
efectos maternos de ambiente permanente; r es el coeficiente
de regresión de orden cúbico para modelar la media de la po- The matrix form of the above complete model, with its
blación, ir y ir son los coeficientes de regresión para modelar respective assumptions, is the following: y Xb Z1a Z2c
los efectos aleatorios directos, genéticos y del ambiente per- Z3m Z4q e; E[y] Xb; E[a] E[c] E[m] E[q]
manente, del r-ésimo polinomio para el i-ésimo animal; ir y E[e] 0. Var(a) ka A; Var(c) kc INd ; Var(m) km
ir, son los coeficientes de regresión para modelar los efectos A; Var(q) kq INm; Var(e) R
aleatorios maternos, genéticos y de ambiente permanente del
r-ésimo polinomio para el i-ésimo animal; y ij , es el efecto where y is the vector of n observations (referring to the number
residual. El orden cúbico para modelar la media de la pobla- of animals with productive records, Nd); b is the vector of
ción () se definió en análisis preliminares considerando sólo fixed effects of contemporaneous groups and coefficients
efectos fijos de año y época de nacimiento y sexo del animal of regression of first, second and third order of the effect of
como clases, más la covariable edad del animal en función li- age; a is a vector, ka ND random regression coefficients for
neal, cuadrática y cúbica. additive direct genetic effects, where ND denotes the total
number of animals in the genetic relationships matrix, with
La forma matricial del modelo anterior completo, con sus res- and without weight records; c is a vector, kc Nd random
pectivas suposiciones es: y Xb Z1a Z2c Z3m Z4q e; regression coefficients for animal permanent environmental
E[y] Xb; E[a] E[c] E[m] E[q] E[e] 0. Var(a) ka effects; m is a vector, km ND random regression coefficients
A; Var(c) kc INd ; Var(m) km A; Var(q) kq INm; for maternal additive genetic effects; q is a vector, kq Nm
Var(e) R random regression coefficients for maternal permanent
environmental effects; Nm is the number of cows that had
donde, y es el vector de n observaciones (referentes al número offspring with data; e is the vector of residual effects; X, Z1,
de animales con registro productivo, Nd ); b es el vector de Z2, Z3, and Z4 are incidence matrixes that relate the vector
efectos fijos de grupos contemporáneos y coeficientes de regre- y to the fixed and random effects described above (b, a, c,
sión de primer, segundo y tercer orden del efecto de la edad; m and q, correspondingly); ka, kc, km and kq are matrixes of
a es un vector, ka ND coeficientes de regresión aleatoria (co)variances of the random regression coefficients for direct
para los efectos genéticos aditivos directos, donde ND denota (additive genetic and permanent environmental) and maternal
el número total de animales en la matriz de parentesco, con y (additive genetic and permanent environmental) effects; A is
sin registro de peso; c es un vector, kc Nd coeficientes de the matrix of additive genetic relationships among the animals
regresión aleatoria para los efectos de ambiente permanente in the pedigree; INd is an identity matrix of dimension Nd ; INm
del animal; m es un vector, km ND coeficientes de regresión is an identity matrix of dimension Nm; Kronecker product;
aleatoria para los efectos genéticos aditivos maternos; q es un and R is a diagonal matrix of residual variances.
328 VOLUMEN 45, NÚMERO 3](https://image.slidesharecdn.com/agrociencia2011tropicarne-120513155316-phpapp01/85/AJUSTE-DE-MODELOS-DE-REGRESION-ALEATORIA-EN-EVALUACIONES-GENETICAS-DE-BOVINOS-TROPICARNE-4-320.jpg)