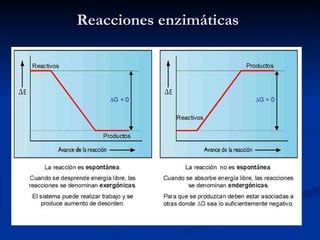
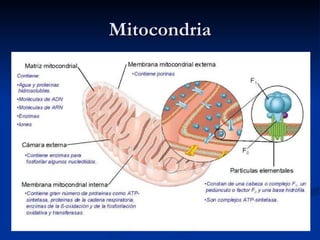
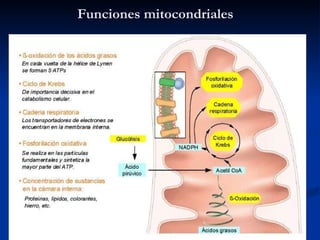
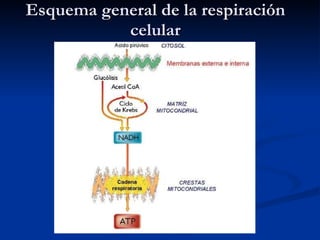
El documento describe los procesos catabólicos aerobios en la célula. La glucólisis y otras rutas degradan moléculas como la glucosa y ácidos grasos en acetil CoA, que alimenta el ciclo de Krebs. Este y la cadena respiratoria en la mitocondria producen electrones que se usan en la fosforilación oxidativa para sintetizar hasta 36 moléculas de ATP por molécula de glucosa inicial.