Descargado 81 veces







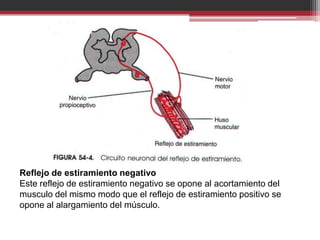



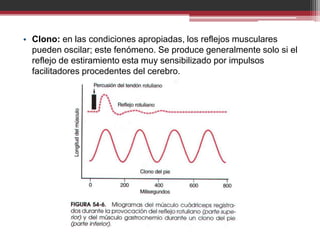








La organización de la médula espinal transmite señales sensitivas y motoras. Las señales sensitivas ingresan por las raíces posteriores y se transmiten a través de interneuronas y motoneuronas anteriores que controlan la contracción muscular. Los husos musculares y órganos tendinosos de Golgi envían retroalimentación sobre la longitud y tensión muscular para regular los reflejos de estiramiento que controlan el tono y movimiento muscular.









































































