Descargado 28 veces






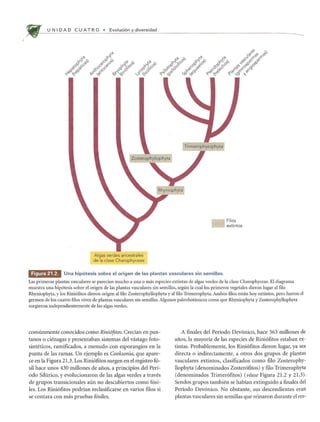






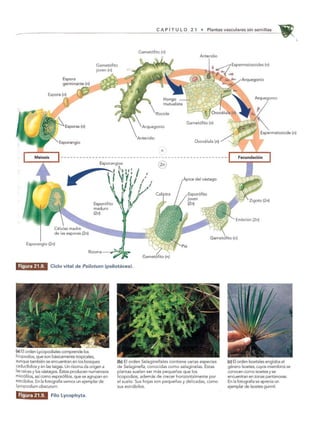


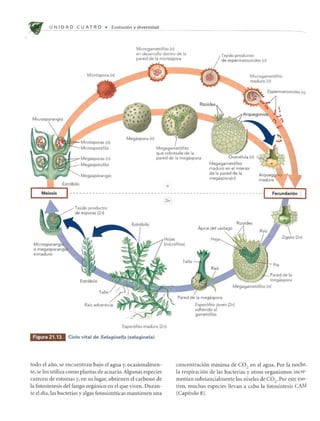



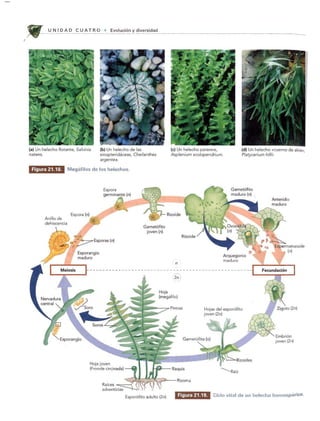





Las plantas vasculares sin semillas dominaban el paisaje hace unos 350 millones de años. Evolucionaron a partir de las algas verdes de la clase Charophyceae hace unos 430 millones de años. Incluían Briófitos, helechos y plantas similares a árboles que crecían en bosques tropicales pantanosos durante el período Carbonífero.
































![Hábito y hábitat [modo de compatibilidad]](https://cdn.slidesharecdn.com/ss_thumbnails/habitoyhabitatmododecompatibilidad-110622195317-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)
![Ensayo de ciencias(completo)[1]](https://cdn.slidesharecdn.com/ss_thumbnails/ensayodecienciascompleto1-101119085639-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)





















































