Descargado 73 veces




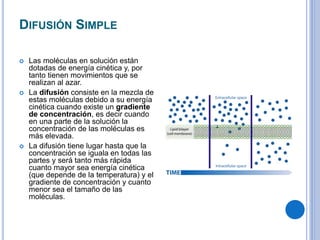

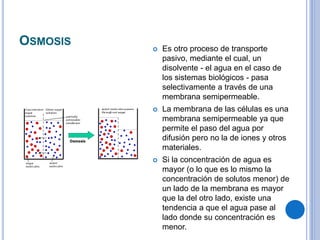
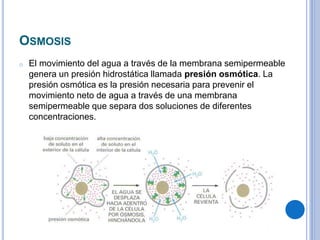
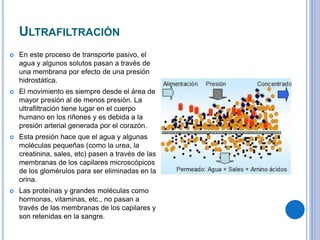
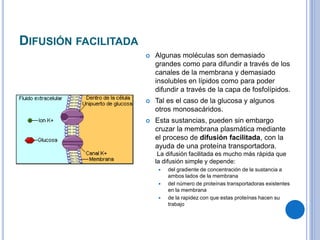


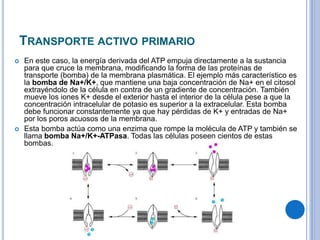
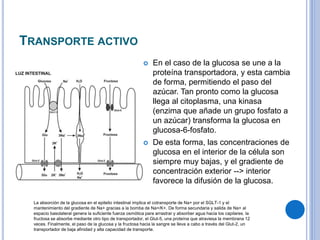

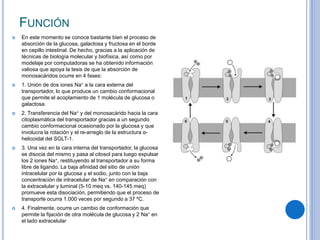



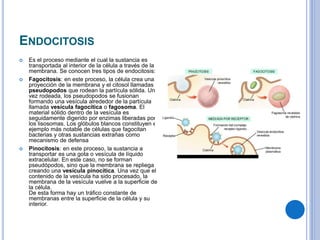

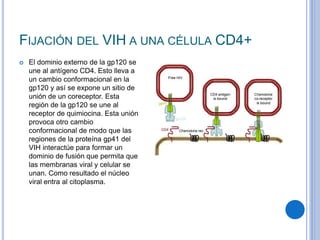
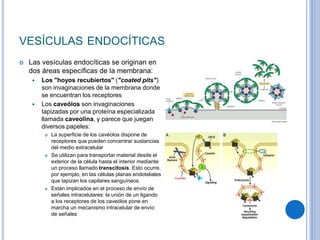
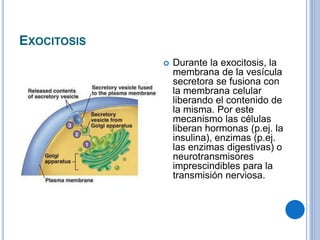

El documento describe los mecanismos de transporte a través de las membranas plasmáticas, incluyendo el transporte pasivo a través de la difusión simple, osmosis, ultrafiltración y difusión facilitada, así como el transporte activo que requiere energía en forma de ATP. Específicamente, se detalla el transporte de glucosa a través de la membrana del intestino delgado usando el transportador SGLT-1 que cotransporta glucosa e iones de sodio.














![Transporte Activo[1]](https://cdn.slidesharecdn.com/ss_thumbnails/transporteactivo1-100313141017-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)








































































