




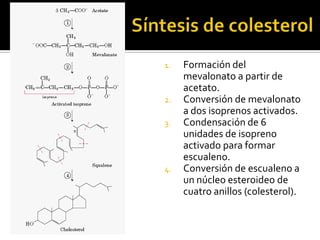

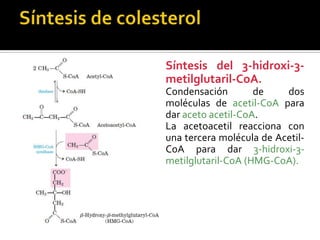
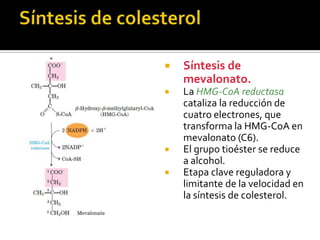
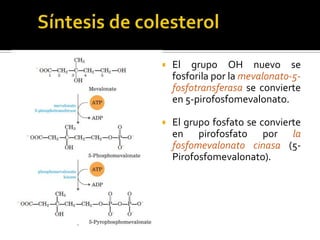
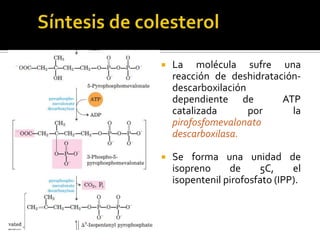
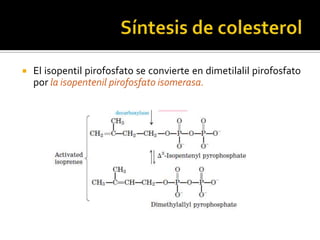

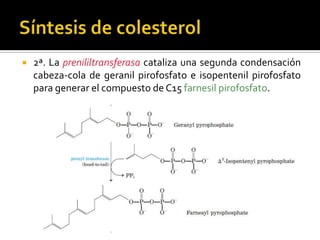
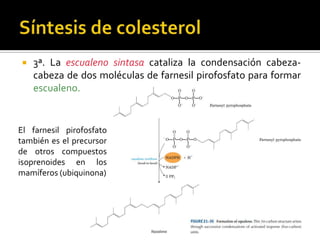
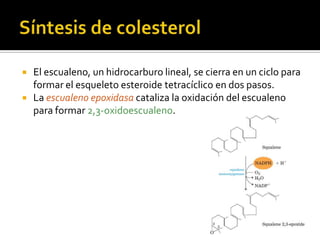
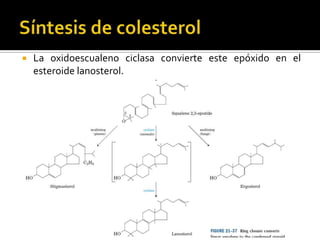
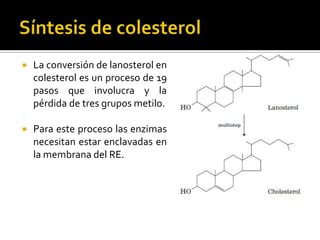
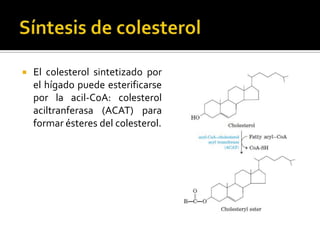


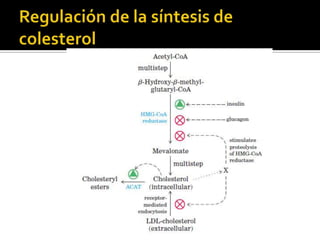
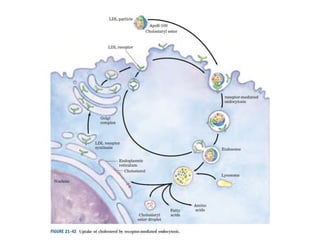
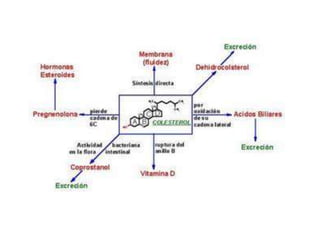



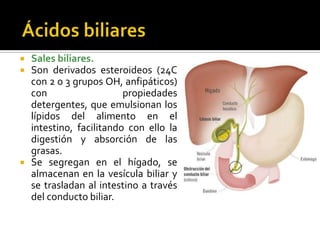

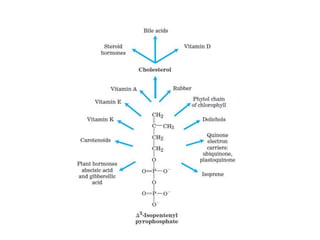


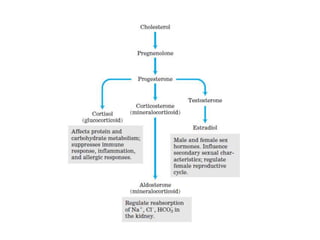


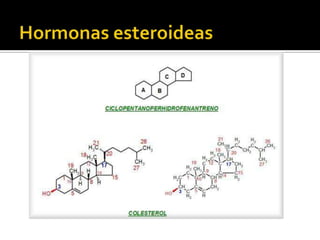
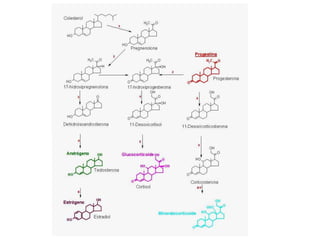


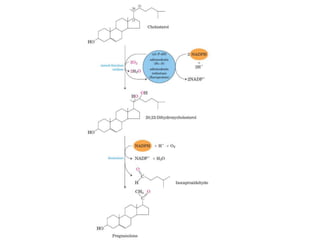




El documento describe la síntesis del colesterol en el cuerpo humano. Se sintetiza principalmente en el hígado, intestino, corteza suprarrenal y tejidos reproductores a partir de acetato. La etapa clave es catalizada por la enzima HMG-CoA reductasa y las estatinas inhiben esta enzima para tratar la hipercolesterolemia. El colesterol es el precursor de las hormonas esteroideas como los glucocorticoides, mineralocorticoides, andrógenos y estrógenos.































































































