ByG 1ºBach Uni9 tejidos y órganos vegetales
•Descargar como PPT, PDF•
2 recomendaciones•11,612 vistas

tejidos y órganos vegetales

Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Destacado (20)
Similar a ByG 1ºBach Uni9 tejidos y órganos vegetales
Similar a ByG 1ºBach Uni9 tejidos y órganos vegetales (20)
Más de VidalBanez
Más de VidalBanez (20)
Último
Último (20)
ByG 1ºBach Uni9 tejidos y órganos vegetales
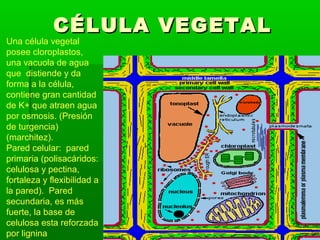
- 1. CÉLULA VEGETAL Una célula vegetal posee cloroplastos, una vacuola de agua que distiende y da forma a la célula, contiene gran cantidad de K+ que atraen agua por osmosis. (Presión de turgencia) (marchitez). Pared celular: pared primaria (polisacáridos: celulosa y pectina, fortaleza y flexibilidad a la pared). Pared secundaria, es más fuerte, la base de celulosa esta reforzada por lignina
- 2. Unidad 9 ORGANIZACIÓN DE LAS PLANTAS: TEJIDOS Y ÓRGANOS VEGETALES Esta presentación se ha elaborado, en proporción variable, a partir de material propio, de otras presentaciones descargadas de la red, y de mi alumnado, actual o pasado. Gracias por su colaboración, a veces desconocida, pero el uso de esta información es puramente educativo. A mi actual alumnado disculpad de cierto exceso en diapositivas con fotos o ilustraciones. Las cambiaré... algún día.
- 4. Tejidos vegetales Concepto de tejido Tipos de tejidos Tejidos embrionarios Tejidos adultos
- 5. Histología Rama del saber que estudia los tejidos Tejido Conjunto de células estructural y funcionalmente característico. Observa la semejanza de las células dentro de cada tipo de tejido.
- 6. Tipos Pueden poseer uno (tejidos simples) o varios tipos de células (tejidos complejos) Tipos Meristemáticos Adultos Basado en: Capacidad de división Diferenciación
- 7. Meristemo s Células vivas, poco diferenciadas Capaces de dar lugar otro tipo de células, tejidos y órganos Meristemo de un tallo Fotografí a de yemas
- 8. Meristemas: Localización Apicales Extremos de los tallos y raíces Crecimiento en longitud Laterales: Cámbium Por toda la planta Responsable crecimiento secundario (“grosor”) Otros: Intercalares, Secundarios
- 9. Tejidos adultos Tejidos simples, un solo tipo de células Parénquima Colénquima Esclerénquima Tejidos complejos formados por varios tipos de células Epidermis Peridermis Xilema Floema
- 10. Xilema Función de transporte de Savia bruta Función de sostén Traqueas Diferentes tipos de células Traqueidas Tráqueas Punteaduras y poros Ambas (muertas y lignificadas) Sólo quedan paredes Dispuestas en alineaciones + Colénquima Esclerénquima Parénquima Traqueidas Sólo punteaduras
- 11. Floema Angiospermas Gimnospermas Elto tubo criboso Cél. cribosa Cél. anexa Cél. albuminosa Células vivas, longitudinales forman los tubos cribosos Pueden carecer de núcleo Transporte de savia elaborada (Hay diferencias entre gimnospermas y angiospermas) + Parénquima Fibras Aspecto del floema en el microscopio electrónico
- 12. Histología Floema Sección longitudinal Angiosperma Sección transversal Angiosperma Elemento de tubos cribosos Célula acompañante Placa cribosa
- 13. Sistemas de tejidos Dérmico Tejidos Epidermis Células Epidérmicas, oclusivas, tricomas Peridermis Parénquima Parenquimáticas Colénquima Fundamental Suberógenas Parenquimáticas Colenquimáticas Fibras Esclereidas Esclerénquima Vascular Xilema Traqueidas Tráqueas Angiospermas / Gimnospermas Floema Elto tubo criboso/ Cél. cribosa Cél. anexa / Cél. albuminosa
- 14. SISTEMA DÉRMICO: cobertura exterior en contacto con el ambiente. Protege de la pérdida de agua, del daño e invasiones de bacterias, hongos e insectos. Hay dos tipos: EPIDERMIS Formada por una o varias capas de células vivas, en función del grado de adaptación a la sequedad. Células aplanadas, con forma a menudo irregulares, interdigitales, ó con formas regulares, poligonales (hexagonales). No presenta espacios intercelulares, bien trabadas Recubre y protege a las estructuras primarias de todo tipo de plantas, sean herbáceas o leñosas (peridermis).
- 15. EPIDERMIS Se cubre de una capa hidrófoba que evita la deshidratación, la cutícula, cuyo componente químico es la cutina. La cutícula es más o menos gruesa en función de esta misma adaptación y del lugar de la planta que cubra.
- 16. EPIDERMIS Las plantas adaptadas a climas secos (xerófitas) acumulan por encima de la cutícula cantidades más o menos notorias de ceras (cera epicuticular) que pueden formar relieves peculiares.
- 21. EPIDERMIS : ESTOMAS Microscopía electrónica de Barrido
- 22. Tejido peridérmico: Reemplaza a la epidermis en tallos y raíces de plantas leñosas adultas. Se compone de células suberosas (corcho) con paredes gruesas e impermeables, que mueren al madurar.
- 23. Peridermis Aparece tras crecimiento secundario Tejido suberógeno (muerto) Cámbium suberógeno (meristema) Felodermis (parenquimático vivo)
- 25. SISTEMA FUNDAMENTAL: células del parénquima, colenquima y esclerenquima. Apoyan, almacenan y secretan. Médula. Corteza. PARENQUIMA Tejido simple formado por células que se encuentran en todo el cuerpo vegetal. Sus células realizan varias funciones según el lugar en que se encuentren. por ejemplo parénquima aerifero en tallos acuáticos Son células redondeadas y de paredes delgadas. Se caracteriza por la presencia de espacios aéreos intercelulares que varían de tamaño. Cumple una función tanto de producción como almacenamiento de alimentos.
- 26. Parénquima fotosintético ó clorofiliano: Se localiza en las partes expuestas a la luz. Poseen cloroplastos encargados de llevar a cabo la fotosíntesis. Parénquima no fotosintético ó aclorofiliano: Se localiza en las partes no expuestas a la luz: las capas profundas de los tallos y en las raíces. Sus células presentan leucoplastos y amiloplastos donde se almacenan sustancia de reserva. PARENQUIMA: TIPOS
- 27. Epidermis Parénquima en empalizada Parénquima lagunar
- 29. Formado por células vivas. Son células alargadas, que presentan paredes secundarias, engrosadas especialmente en las esquinas. Se localizan en las regiones de la planta en crecimiento a fin de darles fortaleza. (zonas jóvenes) Se localizan en la capa más externa de la corteza de los tallos. COLÉNQUIMA
- 30. • Colénquima: consiste en células alargadas poligonales con paredes de espesor irregular, están vivas en la madurez pero generalmente no pueden dividirse, las paredes celulares son fuertes y flexibles. Sirve de soporte en hierbas y pedúnculos
- 32. Esclerénquima Consiste en células, ya muertas, con paredes celulares secundarias gruesas y endurecidas, reforzadas por la lignina. Dan sostén y fortaleza al cuerpo de la planta, pero mueren luego de diferenciarse (fibras del cáñamo y yute, cáscara de nueces, etc.). Generalmente tejidos maduros
- 34. SISTEMA VASCULAR: transportan fluidos y sostienen la planta en posición vertical. Xilema: lleva agua y minerales de las raíces al vástago por tubos hechos de traqueidas y elementos de los vasos.
- 35. • El paso final de la diferenciación de ambos es la muerte: el citoplasma y la membrana celular se desintegran, dejando un tubo hueco de pared celular
- 36. • Floema: lleva agua con sustancias disueltas sintetizadas por la planta, como azúcares, aminoácidos y hormonas. Está formado por células esclerenquimatosas, elementos de tubo criboso y células acompañantes. Al madurar los elementos, se desintegran dejando solo una capa delgada de citoplasma que reviste la membrana plasmática
- 37. En los extremos de los elementos del tubo criboso, donde se unen células adyacentes, se forman agujeros en la pared celular primaria para crear placas cribosas. Las células acompañantes nutren a los elementos a través de plasmodesmos.
- 38. Tejidos: Tallo Colénquima Parénquima Cutícula Epidermis Histología Radios parenquimatosos (xilema)
- 39. Histología Tejidos: Hoja gramínea Esclerénquima Parénquima Xilema Epidermis Floema
- 40. La raíz Concepto Funciones básicas Estructura primaria Estructura longitudinal Modificaciones ¿podrías responder a algo de lo anterior?
- 41. Funciones de la raíz Raíz Obtención de agua y nutrientes Circulación Savias Sujeción Almacén de reservas Órgano generalmente subterráneo y con geotropismo positivo
- 42. Sistema de raíces Raíz Raíz axonomorfa: Predominio raíz principal Raíz fasciculada: No hay predominio raíz principal Raíz primaria: Origen radicular Raíz adventicia: Origen caulinar
- 46. Clases de Raíces y Adaptaciones 5 46
- 47. Tipos de raíces Raíces fúlcreas Raíces adherentes ¿FUNCIÓN de cada tipo? Pneumatóforos y Zancos Raíces tabulares
- 48. Tipos de raíces Raíces reserva Zanahoria, remolacha Raíz
- 50. Tallo Concepto y funciones Definición Transporte Sostén Elevar aparato fotosintético Almacen
- 51. Estructura longitudinal Tallo Yema terminal Yema lateral Cicatriz de un escama de yema Cicatriz de una hoja Nudo Entrenudo Tallo lateral Nudo Cicatriz de un escama de yema Yema lateral
- 52. Partes del tallo ¿Cuál ES CUAL? Tallo principal Tallo secundario Nudo Entrenudo Yema secundaria Yema axilar Yema principal
- 53. Modificaciones del tallo Propagación lateral Rizomas Estolones Tallo
- 54. Tallo Modificaciones del tallo Reserva Tubérculos caulinares Rizoma Tubérculo caulinar: patata Rizoma: Iris
- 55. Modificaciones del tallo Tallo Reserva Bulbos Cormos Tunicado: cebolla No tunicado: Lilium Cormo: Gladiolo
- 60. Hoja Órgano fotosintético, generalmente aplanadas, que surgen del tallo. Generalmente tienen crecimiento limitado Respiración Transpiración Modificaciones para otras funciones Gran variabilidad morfológica
- 61. Partes de la hoja Ápice Limbo Márgen Hoja Base de la hoja Peciolo Limbo o lámina Estípula Estípulas Vain a
- 62. Coloca el nombre donde corresponda . vaina . nervadura . base . peciolo . yema axilar . borde . ápice . envés . limbo . haz
- 63. Nerviación Hoja Dicótoma Nervio medio o principal Palmatinervia Paralelinervia Pinnatinervia
- 64. Hojas simples o compuestas Hoja Limbo entero: simple Limbo dividido: compuesta Truco: En las axilas de las hojas hay yemas, en las axilas de los foliolos no hay yemas Simple Compuesta Foliolo Peciolulo Raquis Axila foliar
- 66. Filotaxis Disposición de las hojas en el tallo Verticiladas Alternas Opuestas Tallo
- 68. Hoja Márgenes Crenada Entera Incisa Serrada Sinuada Serrulada Undulate Lobada Doble serrada Dentada
- 69. Ápice Acuminado Agudo Mucronado Agudo Auriculado Cordado Oblicuo Redondeado Sagitado Hastado Truncado Cuspidado Emarginado Obcordado Base foliar Obtuso Truncado
- 71. Modificaciones de la hoja Cotiledones Filodios
- 72. Modificaciones de la hoja Zarcillos Almacén de agua
- 73. Modificaciones de la hoja Hojas carnívoras
- 74. Modificaciones de la hoja Reserva: Peciolos engrosados Reserva: Bulbos Cebolla Acelga
- 75. Estructura de una hoja limbo nervios cutina epidermis xilema parénquima en empalizada tubos conductores espacio intercelular parénquima lagunar estoma floema
- 76. HOJA: Estructura interna Sección transversal Epidermis Parénquima en empalizada Colénquima Mesófilo Parénquima esponjoso Xilema Haz vascular Floema Epidermis inferior
- 77. Estructura interna Cutícula Epidermis múltiple Hoja xerófila Parénquima en empalizada Nervio con xilema y floema Parénquima esponjoso Estoma Tricoma Adelfa (Nerium oleander)
- 78. Estructura primaria Esclerénquima Hoja Gramínea C3 Parénquima Xilema Células bulliformes Epidermis superior Floema
- 80. Estructura interna Hoja Mesofilo Endodermis Conducto resinífero Xilema Floema Tejido de transfusión Acícula de pino Hipodermis Epidermis Estomas hundidos
- 82. Meristemos apicales: están situados en los extremos de las raíces y los vástagos, lo que incluye los tallos principales y ramas. ► Meristemos laterales: también llamados cambia (cambium), forman cilindros que corren paralelos al eje longitudinal de las raíces y los tallos. ► Clases de Meristemos
- 83. Raíz, Estructura primaria Epidermis: Incluye pelos radiculares. Córtex: Tejido parenquimático y endodermis con banda de Caspary (impermeable) Cilindro vascular: Xilema, floema, periciclo (parénquima) Sección transversal
- 84. raíz
- 85. Raíz, Estructura primaria Sección transversal Floema Xilema Haz vascular Xilema Periciclo Endodermis (con banda Caspary) Procámbium Rizodermis Córtex
- 86. Raíz, Sección transversal de una monocotiledónea (médula) xilema
- 87. ¿Cuál ES UNA RAIZ DE MONOCOTILEDONEA Y CUAL DE DICOTILEDONEA?
- 88. Tallo, Estructura primaria Sección transversal Haces vasculares Epidermis Córtex Haces vasculares Médula Epidermis Médula Xsection de un tallo Córtex Medicago sativa
- 89. Tallo, Estructura primaria Sección transversal Haces vasculares Epidermis Córtex Haces vasculares Médula Epidermis Médula Córtex Tilia americana
- 90. Haces vasculares Estructura primaria Tallo Sección transversal Epidermis Floema Xilema Vaina de esclerénquima Parénquima Monocotiledónea Maíz (Zea mays)
- 91. Tallo
- 92. Concepto Crecimiento secundario Desarrollo en grosor de los vegetales debido a la acción de meristemas laterales Ocurre en tallos y raíces No en todas las especies Como resultado aparece la madera Cámbium vascular Cámbium suberógeno Crecimiento vascular Crecimiento de peridermis
- 93. Formas de Crecimiento • Crecimiento primario: permite el aumento en longitud y el desarrollo de estructuras especializadas. • Crecimiento secundario: causa el aumento en diámetro de tallos y raíces.
- 94. Interior tronco Célula del cámbium Crecimiento secundario Radio parenquimático Xilema secundario Radio inicial parenquimático Cámbium bifacial Floema secundario Exterior tronco
- 95. Crecimiento secundario Tallo Médula Xilema 1º año Xilema Cámbium 2º año Tejido vascular Radio floemático dilatado Epidermis Corteza (Felodermis) Floema secundario Tilia americana 2 años
- 96. Anillos de crecimiento En climas estacionales Crecimiento discontinuo del cámbium Coronas circulares en la madera Anillos crecimiento ¿Número de anillos = Edad árbol? Ojo: Anillos ausentes, falsos anillos Mucha más información: crecimiento, clima, historia, edad Dendrocronología Madera
- 97. Peridermis Cámbium suberógeno o felógeno Felodermis hacia el interior (parenquimático vivo) Súber hacia el exterior, células suberificadas y aislantes Crecimiento secundario
- 98. Lenticelas Aperturas en el súber Difusión de oxígeno Diferentes formas Sambucus
- 99. Aspecto externo Crecimiento secundario El modo en que se desarrolla nueva felodermis: Aspecto de la corteza externa
- 100. Crecimiento secundario Corteza Corteza: Todos los tejidos al exterior del cámbium vascular Corteza exterior Corteza interior Cámbium vascular Xilema secundario Aumento de tamaño– incremento de presión Periódicamente agrietamiento y rotura Desarrollo de nuevo felógeno y nueva felodermis, más al interior
- 101. Madera Duramen y albura Parte funcional: Albura (transporte) Parte no funcional: Duramen (sostén) Corteza secundaria Albura Duramen
- 103. Partes de la Semilla • A) embrión 1) radícula 2) plúmula 3) hipocotilo 4) cotiledón B) endospermo C) epispermo epispermo(5) Micrópilo (6)
Notas del editor
- Tema 18: Células y Tejidos vegetales: Concepto de histología. Tipos de tejidos y sus células constituyentes. Tejidos embrionarios: Meristemas. Tejidos adultos: parénquima, colénquima, esclerénquima, xilema, floema, epidermis y peridermis. Sistemas de tejidos: fundamental, vascular y epidérmico.
- La rama del saber que estudia los tejidos se denomina histología. Un tejido puede definirse como un conjunto de células estructural y/o funcionalmente característico. Los tejidos pueden estar formados por un único tipo de célula (simples) o por varios tipos (complejos).
- Una primera división en dos tipos de tejidos distingue los tejidos embrionarios o meristemas y los tejidos permanentes o adultos. La diferencia entre ambos se basa en que mientras la división celular es característica de los meristemas, este es un proceso raro en los tejidos permanentes. Por otra parte los meristemas muestran una menor diferenciación pudiendo dar lugar a otro tipo tejidos.
- Durante la fase embrionaria las células muestran una gran capacidad de diferenciación, pero conforme alcanzan un cierto grado de diferenciación muchas de ellas pierden la capacidad de dar otro tipo de células. Los tejidos meristemáticos (meristemas) son un tejido formado por células poco diferenciadas que mantienen la capacidad de poder transformarse en diferentes células y tejidos.
- Se disponen en diferentes partes de la planta. Los meristemas apicales se disponen en los extremos de los tallos y raíces y son los responsables del crecimiento en longitud. Los meristemas apicales del tallo suelen estar protegidos por una serie de hojas involucrales, conformando las yemas. Los meristemas laterales se extienden por toda la longitud de la planta formando un cilindro, siendo los responsables del crecimiento secundario de tallo y raíz. En algunas plantas también existen meristemas intercalares que difieren de los meristemas apicales, por estar dispuestos entre tejidos más o menos diferenciados, generalmente en la base de los entrenudos. Pueden aparecer meristemas secundarios a partir de células diferenciadas, indicando que éstas no han perdido totalmente su capacidad de diferenciarse.
- El xilema es el responsable de la conducción de la savia bruta a través de la planta. Además de esta función, tiene una importante labor estructural junto a colénquima y esclerénquima. Se puede diferenciar el xilema propio de la mayor parte de las gimnospermas con las traqueidas como único elemento de conducción, del poseído por la práctica totalidad de las angiospermas, que cuenta además con otro tipo de células, las tráqueas. Tanto tráqueas como traqueidas, tienen en común estar muertas en su madurez, quedando huecas y con una pared secundaria lignificada. En ambos casos se disponen formando largas alineaciones que comunican la raíz con las diferentes partes fotosintéticas de la planta. Las traqueidas tienen una forma más alargada y acabada en punta que las tráqueas. Sin embargo, su diferencia principal estriba en que las tráqueas poseen punteaduras y perforaciones en su pared, mientras que las traqueidas únicamente poseen punteaduras, por lo que el paso de solutos es algo menos eficiente. El tejido xilemático además de este tipo de células posee zonas de parénquima, colénquima o fibras de esclerénquima.
- El floema es el encargado del transporte de la savia elaborada. De igual modo que el xilema, sus elementos principales son células elongadas que forman estructuras longitudinales. A diferencia del xilema, estas células están vivas en la madurez, aunque puedan carecer de núcleo.
- Las células conductoras poseen una zona de contacto denominada placa cribosa con poros para comunicarlas, además están acompañados por otras células adyacentes, cuya función es regular el flujo de salida y entrada de nutrientes al sistema. Existe también una diferenciación entre los tejidos floemáticos propios de las gimnospermas y aquellos propios de las angiospermas. En las gimnospermas estas funciones corresponden a la célula cribosa y a una célula parenquimática modificada, la célula albuminífera, que provienen de diferentes células madres. En el caso de las angiospermas, aparecen dos células: elemento de los tubos cribosos y células acompañantes, ambas provenientes de una misma célula madre. Del mismo modo que en el xilema, en el floema pueden aparecer células parenquimáticas, así como fibras o esclereidas.
- La epidermis es la capa de células que rodea el cuerpo del vegetal en todos sus órganos. La epidermis está formada por células vivas densamente trabadas entre sí, estas células generalmente se disponen en una única capa, pero en algunos casos especiales pueden haber varias capas. Generalmente, sus paredes están recubiertas exteriormente por una capa de cutícula (principalmente cutina y ceras) que las impermeabiliza del exterior. Además de estas células, la epidermis tiene distribuidas por su superficie una serie de células especializadas, poseedoras de cloroplastos, que son las que se encargan de regular el paso de los gases (células estomáticas). Estas células suelen estar asociadas a células epidérmicas que regulan su apertura. Un aspecto muy importante en las epidermis es la habitual presencia de diversos pelos y tricomas con múltiples funciones (secretoras, protectoras, absorbentes) que pueden ser uni o pluricelulares.
- Cuando los órganos de las plantas sufren fuerte crecimiento secundario la epidermis es sustituido por la peridermis. La peridermis cuenta con tejido suberógeno muerto en el exterior, una capa de cambium suberógeno y un tejido parenquimático en el interior (felodermis). El cámbium genera ambos tejidos por división. Esta estructura es la que conforma las cortezas de los árboles.
- Cuando los órganos de las plantas sufren fuerte crecimiento secundario la epidermis es sustituido por la peridermis. La peridermis cuenta con tejido suberógeno muerto en el exterior, una capa de cambium suberógeno y un tejido parenquimático en el interior (felodermis). El cámbium genera ambos tejidos por división. Esta estructura es la que conforma las cortezas de los árboles.
- El parénquima está formado por un tipo de células, las células parenquimáticas. Una de las propiedades principales de las células parenquimáticas es que están vivas. Estas células generalmente son isodiamétricas y rara vez están elongadas. El parénquima a menudo se encuentra rellenando los huecos entre otras células más especializadas. Las células parenquimáticas no están muy diferenciadas y no es raro que posean cloroplastos o que puedan dar lugar a otros tipos de células. Los parénquimas pueden ser de diferentes tipos y cumplir diferentes funciones: tejido fotosintético, almacenamiento de substancias, elemento de transferencia, almacenamiento de líquidos y espacios para el paso del aire (aerénquima). Las células parenquimatosas tienen la pared celular poco engrosada
- El colénquima está formado por células colenquimáticas. Estas células al igual que las células parenquimáticas se mantienen vivas durante la madurez. El colénquima suele disponerse en forma de haces o cilindros discretos. La función básica del colénquima es servir de sostén del tejido primario en crecimiento. Para en ello en la estructura de sus paredes primarias aparecen zonas de donde se acumula celulosa y pectina, lo que les otorga resistencia y flexibilidad. Parece que el colénquima se puede desarrollar a partir del parénquima.
- El esclerénquima, a diferencia de los tejidos anteriores, está formado por células muertas. En este tejido las células producen gruesas paredes secundarias. Se trata de un tejido de sostén. Existen dos tipos de células que pueden conformar este tejido, las fibras y las esclereidas. Las fibras son largas y delgadas y se suelen agrupar en haces o cordones, muchas de ellas tienen un gran interés económico (lino, pita, cáñamo...). Las esclereidas se pueden presentar aisladas o agrupadas por todo el tejido fundamental (cáscara de la nuez, hueso de la aceituna).
- El esclerénquima, a diferencia de los tejidos anteriores, está formado por células muertas. En este tejido las células producen gruesas paredes secundarias. Se trata de un tejido de sostén. Existen dos tipos de células que pueden conformar este tejido, las fibras y las esclereidas. Las fibras son largas y delgadas y se suelen agrupar en haces o cordones, muchas de ellas tienen un gran interés económico (lino, pita, cáñamo...). Las esclereidas se pueden presentar aisladas o agrupadas por todo el tejido fundamental (cáscara de la nuez, hueso de la aceituna).
- Aspecto de un tallo
- Aspecto de una hoja
- Tema 19: La raíz Concepto. Funciones básicas. Sistemas de raíces. Estructura primaria de la raíz. Estructura longitudinal de la raíz. Modificaciones de la raíz.
- La raíz es un órgano vascularizado, generalmente subterráneo, que suele servir a la planta para obtener agua y nutrientes, así como de sujeción y/o de almacén de reservas.
- Se denomina raíz primaria a la que proviene directamente de la radícula del embrión. En función de la dominancia de la raíz primaria se establecen dos sistemas radicales: sistema axonomorfo donde predomina la raíz principal con raíces laterales de menor importancia y sistema de raíz fasciculada, donde no hay un predominio de la raíz principal y aparecen numerosas raíces de importancia semejante. Las raíces que se originan en los tallos se denominan raíces adventicias.
- La raíz puede tener diferentes orígenes y funciones. Existen raíces de origen caulinar como son las raíces fúlcreas del maíz con origen caulinar y función de sujección. Las raíces que se desarrollan fuera de la tierra se denominan raíces aéreas, este es el caso de las raíces de la hiedra que sirven para fijar el tallo al sustrato sobre el que se desarrolla o como las raíces aéreas de los mangles que tienen como finalidad la respiración (pneumatóforos), otras raíces como las del ciprés de los pantanos (Taxodium distichum) tienen como objetivo conferir mayor estabilidad a la planta
- Muchas raíces sirven para almacenar reservas para las plantas, estas reservas casi siempre se realizan en forma de carbohidratos, generalmenne almidón. Es el caso de las raíces engrosadas de zanahorias, rábanos o remolachas, pero también el de muchas otras plantas como el diente de león. Estos órganos de reserva sirven para que las plantas almacenen el resultado de los años favorables, para ser utilizadas en otros momentos (escasez, floración, herbivorismo). En este sentido cabe destacar el hecho de que numerosas plantas utilizan estas reservas para rebrotar en el caso de ser cortadas o tras los incendios. Algunas raíces como en ciertas orquídeas epifitas pueden realizar la fotosíntesis.
- El vástago está compuesto por el tallo y sus hojas. El tallo es un órgano cuya función principal es levantar y soportar los aparatos fotosintéticos y reproductor, así como comunicarlos con el sistema radicular.
- Los meristemas de crecimiento primario del tallo se sitúan en unas estructuras denominadas yemas. Las yemas pueden aparecer en diferentes lugares, y se distingue generalmente entre las yemas apicales, situadas en el ápice de los tallos y las yemas laterales, situadas en las ramificaciones secundarias. El predominio de uno u otro tipo de yemas, determina la forma de la planta. Si en el crecimiento del tallo se da un predominio de la yema apical, hablamos de un tipo de crecimiento monopódico. En caso de que este predominio se pierda y se desarrollen de modo parecido las yemas laterales hablamos de crecimiento simpódico. Las podas buscan alterar estos equilibrios para favorecer el crecimiento que deseemos. Un análisis de la estructura longitudinal del tallo permite observar que este se estructura de una forma metamérica con zonas sin ramificación (entrenudos) y otras en las que se produce la ramificación (nudos).
- Además de cumplir sus funciones básicas, el tallo o diferentes partes de él pueden sufrir modificaciones para cumplir otros objetivos. Algunos tallos pueden ser subterráneos, incluso ramificándose, (rizomas). Otros se extienden por la superficie (estolones) permitiendo en muchos casos el establecimiento de otros individuos de modo vegetativo.
- El papel de almacenamiento de reservas es común a la mayor parte de los tallos, si bien en algunos casos se desarrollan estructuras especiales, como los tubérculos caulinares formados en estolones (Solanum tuberosum), engrosamiento del epicotilo (Brassica oleracea var. gongylodes), bulbos en los que intervienen también las hojas, siendo en algunos casos las más importantes como en la cebolla (Allium cepa), si bien en otros hay predominio de los tejidos caulinares (Gladiolus sp.).
- Algunos tallos se transforman en estructura defensivas formando espinas (p.ej. Prunus spinosa) o aguijones (Rosa sp.). Los aguijones, a diferencia de las espinas en las que participa todo el tallo, se desarrollan únicamente a partir del córtex y parte de la epidermis.
- En algunos casos los tejidos caulinares sirven de almacenamiento de agua (como en numerosos cactus y euforbias). Otros pueden asumir el papel de órganos fotosintéticos, asemejándose a hojas (cladodios de Ruscus aculeatus).
- La hoja es el órgano sobre el que suele recaer la función fotosintética, generalmente es aplanado. Presenta una gran variedad de formas. De modo secundario puede transformarse para tomar otras labores
- El estudio de la morfología externa de las hojas es fundamental para el reconocimiento de las diferentes especies. Una hoja ideal constaría de una lámina aplanada (limbo) que está unido al tallo por mediante el pecíolo, a menudo en la base del pecíolo pueden aparecer unas estructuras foliáceas (estípulas). En algunas plantas como las gramíneas, pero también en otros grupos, pueden aparecer expansiones foliares que abrazan el tallo (vaina). Todas estas estructuras pueden estar fuertemente modificadas, o incluso faltar algunas de ellas. La parte basal de la hoja por donde se une al peciolo se llama base de la hoja y el extremo opuesto ápice de la hoja, el contorno externo de la hoja es el margen.En caso de que falte el peciolo las hojas se denominan sentadas o sésiles.
- Las nerviación de las hojas tiene diferentes tipos. Si todos los nervios discurren paralelos como es común en las monocotiledóneas y en unos pocos grupos de dicotiledóneas hablamos de nerviación paralela u hojas paralelinervias, Si hay un nervio principal del que salen en diferentes puntos nervios secundarios más cortos hablamos de nerviación pinnada u hojas pinnatinervias. Si aparecen diferentes nervios de orden principal que parten en la base de la hoja en direcciones divergentes hablamos de nerviación palmeada u hojas palmaticompuestas. Un tipo de nerviación bastante raro que ocurre en los helechos y en algunos espermatofitos como el caso de Ginkgo es la nerviación dicótoma en la que cada nervio se divide progresivamente en grupos de dos nervios iguales.
- Cuando el limbo es continuo se considera que son hojas simples, pero si el limbo está divididp enteramente entonces hablamos de hojas compuestas. láminas más pequeñas (foliolos), que se unen al eje foliar (raquis) mediante pequeños pecíolos (peciolulos), son las hojas compuestas. Distinguir una hoja simple de una compuesta, a menudo resulta complicado para el alumno, pero resulta sencillo si tenemos en cuenta que en la unión entre hoja y tallo (axilas foliares) aparece siempre una yema de crecimiento, no así en la unión de los foliolos con el raquis.
- Entre las hojas compuestas podemos distinguir diferentes tipos en función de como están divididas. Aquellas que muestran sus foliolos unidos en un mismo punto se denominan palmaticompuestas, como es el caso del castaño de indias (género Aesculus). Cuando los foliolos se distribuyen en un eje, se denominan pinnaticompuestas como es el caso de la rosa, el fresno, el lentisco o algunas acacias. Si el foliolo terminal es impar se denominan imparipinnadas como las rosas, si el foliolo terminal es par, se denomina paripinnadas. En caso de que las hojas estén pinnadas más de una vez son hojas bipinnadas. Mención aparte merecen las hojas que están en grupos de tres y que se denominan trifoliadas.
- Un aspecto a considerar es la filotaxis de las hojas, esto es, el modo de disponerse a lo largo del tallo. De un modo muy simplificado pueden establecerse tres categorías. Alternas cuando se disponen por el tallo de modo aislado, opuestas cuando las hojas aparecen unidas al tallo a la misma altura en grupos de dos y verticiladas cuando se unen al tallo a la misma altura en número superior a dos.
- Las hojas son enormemente variables en cuanto a su forma y existe una riquísima terminología para describirla, este es un ejemplo de las formas básicas. Su manejo con una cierta soltura es necesario para la correcta determinación de las plantas.
- Al igual que con la forma del limbo existe una importante terminología definiendo el margen del limbo.
- En función de cómo sea el ápice del hoja o la base foliar existe una amplia tipología de las que estas formas son sólo una muestra.
- Las hojas también se diferencian por su longevidad, que puede oscilar entre unos pocos meses y años. Este aspecto es de gran interés para clasificar árboles y arbustos, siendo perennes cuando duran más de un año y caducifolias cuando caen durante a estación desfavorable mediante un proceso denominado abscisión. Las hojas marcescentes son un tipo especial de hojas caducas, que después de secarse son retenidas en el árbol hasta el brote de las nuevas hojas.
- Las primeras hojas que desarrolla una planta y que ya están en la semilla suelen ser diferentes del resto de las hojas, y se denominan cotiledones. En los estadios iniciales los cotiledones suelen tener tejido de reserva con el que nutren al resto de la planta. En algunas plantas los limbos desparecen desarrollándose los peciolos como hojas (filodio).
- Las hojas pueden tener muchos tipos de modificaciones. Algunas se engruesan para poder acumular grandes cantidades de agua, son las hojas crasuláceas. Otras desarrollan zarcillos que les permiten asirse a otras plantas o a elementos inertes y apoyarse en ellos para erguirse.
- En algunos casos las hojas adquieren mecanismos para la captura de animales con el fin de obtener nutrientes que son escasos en los suelos. Los mecanismos pueden ser muy diferentes. Los más sencillos tienen hojas pegajosas como es el caso del género Pinguicula. Pueden desarrollarse pelos glandulosos con substancias pegajosas (Drosera o Drosophylum) a los que el animal queda adherido, segregando posteriormente substancias que los digieren para ser absorbidos por el tejido foliar. En el género Dionea aparecen mecanismos de cierra más o menos rápido, mientras en Nepenthes o en aparecen trampas en forma de bolsa como el género Sarracenia.
- El papel de almacenamiento de reservas es común en muchas de las hojas, recordemos que los bulbos además de tallos incluyen hojas modificadas. Otras plantas acumulan sus reservas en los peciolos de las hojas como ocurre con las acelgas.
- Si consideramos la estructura interna de una hoja. Podemos observar una estructura interna más o menos semejante. Por ambos lados aparecerán sendas capas de epidermis cubiertas de una capa de cutina que evitará su desecación. A su vez en el interior aparece el mesófilo predominará un tejido parenquimático con células fotosintéticas, puntualmente pueden aparecer capas de tejido de sostén (colénquima o esclerénquima). El mesófilo se ve atravesado por los haces vasculares de floema y xilema. Este patrón general tiene una cierta variabilidad dependiendo del tipo de hoja y el ambiente en que se desarrolla la especie en cuestión. En general entre las plantas dicotiledóneas pueden distinguirse claramente dos tipos de parénquimas, uno formado por células apretadas densamente que ocupan la parte inmediatamente bajo el haz de las hojas, mientras que bajo ellas aparece un parénquima con muchos espacios vacíos que facilitan la difusión de los gases, parénquima esponjoso.
- En algunas dicotiledóneas adaptadas a ambientes muy secos, pueden aparecer algunas adpataciones para reducir la pérdida de agua. Este es el caso de la adelfa (Nerium oleander) un arbusto muy común en cursos de agua temporales de ambientes mediterráneos, donde puede haber una intensa sequía estival. Además de la gruesa capa de ceras (cutícula en la superficie de la hoja), la epidermis tiene varias células de espesor, contra la epidermis de una única capa típica de las especies de ambientes más mésicos. Por otra parte, tal y como ocurre en muchas plantas los estomas se concentran en la parte inferior de la hoja, con el fin de reducir la transpiración. Esta parte presenta además unos pequeños pelos (tricomas) Estos estomas aparecen en pliegues interiores de las hojas (criptas estomáticas) para así reducir el gradiente de humedad. El mesófilo es más o menos semejante al otro tipo de hojas con un parénquima en empalizada y un parénquima esponjoso. Todo ello atravesado por los haces vasculares.
- Las hojas de las gramíneas muestran algunas diferencias. En primer lugar es común la presencia de haces de esclerénquima en el mesófilo, lo que les da una estructura más rígida. Generalmente no se aprecia una diferencia dos tejidos parenquimatosos diferenciados. Un aspecto muy llamativo es la presencia de unas células de gran tamaño en la epidermis (células bulliformes), cuya finalidad parece relacionada con el pliegue de las hojas en condiciones de sequía. Los haces vasculares discurren paralelos como corresponde a una monocotiledónea.
- Un caso muy particular son las gramíneas con fotosíntesis C4, como es el caso del maíz (Zea mays). En este tipo de plantas se observa como las células del mesofilo ricas en cloroplastos se disponen en forma de corona (anatomía Kranz) alrededor de los haces vasculares.
- Las hojas de los pinos muestran unas extraordinarias adaptaciones a ambientes áridos y su diseño se separa mucho de los que hemos visto con anterioridad. En primer lugar no hay una división clara entre cara superior y cara inferior de la hoja. Toda la epidermis se haya rodeada por una capa gruesa de cutícula, aparece por ahí una capa exterior la epidermis. Por debajo de ella aparecen varias filas de células compactas (hipodermis). Por debajo aparece el mesofilo, con células parenquimáticas y con cloroplastos. Este mesofilo suele estar atravesado por varios conductos resiníferos que sirven como defensa frente a herbívoros. En el centro de la hoja aparece uno o dos haces vasculares rodeados por un tejido denominado de transfusión. Este tejido está rodeado por una capa celular (endodermis) que lo separa del mesofilo.
- En una sección transversal de la raíz se pueden distinguir tres zonas: a) la epidermis con los pelos radiculares, que son los encargados de la absorción de agua y nutrientes, b) el córtex con los diferentes tipos de tejidos parenquimáticos y una banda interna de endodermis con la banda de Caspary, de carácter impermeable y que la separa de c) el cilindro vascular con los tejidos vasculares y el periciclo.
- Los pelos radiculares son los encargados de captar el agua y los nutrientes del suelo, tienen una vida muy corta y aparecen casi exclusivamente en la zona de maduración de la raíz. Dentro del córtex predomina el tejido parenquimático con espacios de aire, estas células suelen almacenar almidón y substancias de reserva. La parte más interna del córtex se denomina endodermis, carece de espacios aeríferos y aparece un capa de suberina (substancia impermeable) entre sus paredes, lo que obliga al agua a pasar a través de las células. En el haz vascular suele aparecer una parte central donde se encuentran los haces de xilema, externamente a estos aparece el floema y en una zona más externa el periciclo que está formado por células parenquimáticas pero que tienen capacidad de dar lugar a nuevos tejidos
- En las monocotiledóneas las raíces suelen tener una estructura diferente en lo que se refiere al haz vascular, tal y como puede observarse en esta imagen.
- La estructura primaria de todos los tallos sigue un mismo patrón general con tres bandas concéntricas: epidermis, córtex y médula, si bien con diferentes variaciones. El córtex incluye los haces vasculares del floema y del xilema, así como tejido meristemático. La médula está formada por tejido parenquimático. Este patrón puede tener diferentes modificaciones.
- Así en algunas especies como en Tilia (tilo) los haces vasculares forman una banda continua con una capa de meristema (cámbium vascular) dispuesta entre ellas. Bajo la epidermis en la parte exterior de la corteza aparece una banda exterior de colénquima que confiere estructura al tallo y tras ella una capa de parénquima.
- Las monocotiledóneas muestran una estructura muy diferente en sus tallos. La principal diferencia radica en la disposición de los haces vasculares (xilema y floema), mientras que en las dicotiledóneas estos muestran una estructura claramente diferenciada, formando una banda continua (tipo tilo) o discontinua (tipo alfalfa). Estos aparecen repartidos por todo el tallo en las monocotiledóneas. La parte exterior de cada haza corresponde al floema y la interior al xilema, entre ellos aparece un tejido meristemático.
- En muchas especies una vez ha tenido lugar el crecimiento primario no existen modificaciones en la estructura de sus órganos, que podrán en algunos casos elongarse por la actividad de meristemas apicales, pero nunca aumentar en grosor. Sin embargo, en otras especies puede aparecer un crecimiento secundario en grosor por la actividad de meristemas laterales (cámbium). Este crecimiento es característico de los órganos de crecimiento indefinido (raíz y tallo).
- La actividad del cámbium genera hacia el interior xilema secundario y hacia el exterior floema secundario. Esta división se produce de modo paralelo a la corteza del vegetal. En general suele existir una sólo capa celular de cámbium, si bien a veces resulta difícil distinguir entre las células cambiales y las ya divididas pero aún no diferenciadas. Generalmente se desarrolla mucho más xilema secundario que floema secundario.
- El crecimiento en grosor del eje vascular origina que los tejidos situados hacia el exterior del cilindro cambial sufran una fuerte compresión por el crecimiento de estos. Esta presión incluye tanto a los elementos floemáticos más antiguos como a la corteza parenquimática y la epidermis exterior.
- En aquellos climas donde los árboles están sometidos a una fuerte estacionalidad, como es el caso de las latitudes templadas, el crecimiento del cámbium vascular no es continuo, sino que sufre parones durante la época desfavorable. Estos pulsos pueden reconocerse en la sección de la madera como coronas circulares concéntricas de diferente espesor, estos son los llamados anillos de crecimiento. El conteo de los anillos puede permitirnos conocer la edad del árbol, si bien hay que considerar que por diferentes motivos, como sequías repentinas pueden aparecer parones breves de crecimiento con formación de un falso anillo anual, o bien situación muy estresantes que impidan la formación de anillo durante un determinado año (anillos ausentes). La anchura de estos anillos varía de año a año dependiendo de diversas causas, tanto intrínsecas del árbol (especie, edad), como debidas a factores ambientales (temperatura, luz, disponibilidad hídrica). El estudio de los anillos de crecimiento permite por tanto conocer tanto la edad del árbol como las condiciones ambientales en que éste se desarrolló, esta disciplina científica se conoce como dendrocronología.
- Este cámbium suberógeno o felógeno, al igual que el cámbium vascular, desarrolla dos tipos de tejidos: súber hacia el exterior y felodermis hacia el interior. El súber está compuesto de células muertas suberificadas y dispuestas de un modo compacto, lo que le convierte en un tejido protector que aísla el tallo, del agua y aire. La felodermis es un tejido parenquimático de naturaleza semejante a la corteza parenquimática. A este conjunto de tejidos se le suele denominar corteza.
- El carácter impermeable del súber hace necesaria la aparición de estructuras que permitan la difusión del oxígeno al interior del eje de crecimiento, con este fin aparecen periódicamente aperturas en el súber (lenticelas). Estas lenticelas se observan exteriormente como incisiones en la corteza que pueden ser ovaladas, alargadas o circulares. También pueden aparecer en algunos frutos como es el caso de la manzana.
- El modo en que la producción de nuevas felodermis tiene lugar determina el aspecto de las diferentes cortezas externas de los árboles. En este sentido pueden observarse muchas diferencias entre las diferentes especies.
- Como resultado del desarrollo del cambium vascular los tejidos situados en su exterior sufren una constante presión. Esto es especialmente notable en el floema secundario que suele verse aplastado por el continuo desarrollo del cambium. El término corteza se refiere a todos los tejidos exteriores al cámbium, que incluye tanto el floema como la peridermis. A medida que se genera xilema y floema secundario el grosor del tronco va aumentando, las partes exteriores se agrietan. A partir de células parenquimáticas aparecen nuevos meristemas que forman el nuevo cámbium suberógeno (felógeno) y a partir de él una nueva felodermis.
- Con el crecimiento en grosor del xilema secundario, ciertas partes internas del xilema pierden su funcionalidad conductora.
- Las raíces pueden tener crecimiento secundario en grosor, del mismo modo que los tallos. Este crecimiento secundario empieza con la formación del cámbium vascular, esta capa de células meristemáticas, concéntricas. Estas células tienen su origen en el procámbium. Este meristema se divide y da a lugar a tejido vascular secundario (xilema hacia el interior de la raíz y floema hacia el exterior) El periciclo también genera un cámbium (felógeno) que va a dar lugar al crecimiento de la corteza. Los tejidos más extrernos del crecimiento primario (endodermis, córtex y peridermis) se han alargado, fracturado y terminan por desprenderse. Las zonas de la raiz con crecimiento secundario son zonas en las que ya no se da absorción de nutrientes ni formación de raices laterales