La estructura y función de la neurona
•Descargar como PPT, PDF•
0 recomendaciones•5,654 vistas


Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Similar a La estructura y función de la neurona
Similar a La estructura y función de la neurona (20)
Último
Último (20)
La estructura y función de la neurona
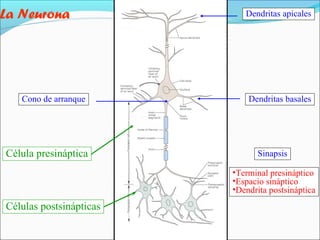
- 1. La Neurona Cono de arranque Célula presináptica Dendritas apicales Dendritas basales Sinapsis •Terminal presináptico •Espacio sináptico •Dendrita postsináptica Células postsinápticas
- 2. Una clasificación neuronal Según el número de prolongaciones del cuerpo celular
- 3. Membrana, Canales e Iones
- 4. Bicapa de fosfolípidos Cabeza hidrófila Cola hidrófoba BCP-F3.3
- 5. Para que un ión se traslade desde su entorno acuoso hasta el interior (donde están las colas de la doble capa, a las que no les gusta el agua) se le debe proporcionar gran cantidad de energía que le permita vencer las fuerzas de atracción entre él y las moléculas de agua circundantes: Por ello la capa es completamente impermeable a los iones. Interior: colas hidrófobas Exterior: cabeza hidrófila
- 6. Estructura de los canales Se requieren 4-6 moléculas de polipéptidos para formar un poro
- 7. Célula K + Aniones Na+ Cl– Ca++ Medio extracelular Sodio: Na+ Cloro:Cl – Potasio: K+ Calcio: Ca++
- 8. Concentración de iones en el axón gigante del calamar Los valores de estas concentraciones en vertebrados son un factor 2 o 3 menor, sin embargo la relación entre ellas es similar.
- 9. Potencial de membrana La membrana bloquea la difusión de iones, lo que da lugar a una diferencia de potencial: V = V −V m in out V V in out Si se abren canales, los iones pueden pasar y afectan a Vm: la entrada de cationes o salida de aniones despolarizan la neurona La salida de cationes o la entrada de aniones la hiperpolarizan
- 10. Potencial de reposo de la membrana V rest Potencial de reposo de la membrana, toma valores tales entre -60mV y –70mV
- 11. La membrana es un condensador KSJ
- 12. Tipos de canales 1. Pueden ser selectivos al tipo de ión: canales de K+, Na+, de Ca2+, ... 2. Pueden ser activos o pasivos 3. Los activos difieren en el mecanismo de activación: Un canal se abre o cierra siguiendo alteraciones de diverso tipo que sufre su microentorno.
- 13. Canal pasivo
- 14. despolarización Cerrado: cerrado y activable Activo: abierto Canal de Na+ activado por voltaje repolarización Refractario: cerrado e inactivable
- 15. Canales activados por ligando (neurotransmisores) Activación directa Activación indirecta
- 16. Canal activado por acetilcolina (Ach)
- 17. Canales activados por glutamato receptor AMPA receptor NMDA activación indirecta
- 19. Regiones Funcionales de una neurona: Transformación de la señal Normalmente poseen 4 regiones funcionales a través de las cuales transforman la señal: de recepción o entrada de activación de conducción de emisión
- 20. Regiones Funcionales La neurona transforma la señal
- 21. 1: Etapa de entrada (input) Neurona sensorial: la señal se transforma en un potencial receptor Interneurona: la señal se transforma en un potencial postsináptico
- 22. Neurona sensorial – Potencial Receptor Transduce la señal original (estímulo físico) en actividad eléctrica Unión neuromuscular: un estiramiento del músculo abre el poro de los canales iónicos, esto altera el potencial de reposo de la membrana. La alteración del potencial es el potencial receptor, que es la primer representación del estiramiento (señal original) Este potencial es típicamente despolarizante (no en la retina)
- 24. Son graduados: su amplitud es función del estiramiento (receptor) o de la cantidad de neurotransmisor recibido (interneurona) Son señales locales: el mecanismo que las genera no es capaz de propagarlas. Su amplitud decrece mucho luego de 1 o 2 mm. El potencial receptor es típicamente despolarizante, los fotoreceptores son un contra ejemplo. El potencial sináptico puede ser Despolarizante: se dice que la sinapsis es excitadora Hiperpolarizante: se dice que la sinapsis es inhibidora El efecto sobre la célula postsináptica depende del tipo de molécula receptora.
- 25. 2y3 - Integración y Potencial de Acción (PA) Cono de arranque (axon hillock) PA o espiga
- 26. Integración y PA
- 27. Transformación de la señal en un código de frecuencia El PA codifica la señal en términos de la frecuencia de disparo: Al crecer la amplitud de la señal aumenta la frecuencia de disparo Al crecer la duración de la señal aumenta el tiempo en que se producen espigas
- 28. Señales locales y propagadas Amplitud Sumación Duración Efecto de la señal Modo de propagación
- 29. Ejemplo Ejemplo de las 4 funciones: neurona sensorial
- 30. Transformación de la señal: resumen
- 31. Un reflejo monosináptico: el Reflejo Patelar (extensión de la rodilla) Un golpe en la rodilla estira al cuádriceps. Esto desencadena un proceso que produce la contracción del cuádriceps y la relajación simultánea del bíceps. La pierna se extiende.
- 33. Inhibición Elimina la actividad de las vías oponentes Amoritgua la actividad del mismo sistema
- 34. Secuencia de señales en la acción refleja
Notas del editor
- El PSP es de la fig KSJesp-F13.3 Texto de KSJesp (pag 35): las neuronas motoras y las interneuronas (ver figura en transparencia anterior) aparece una señal de entrada cuando la neurona presináptica libera neurotransmisor.
- KSJ pag 126 Cuando la despolarización llega a un cierto nivel (valor umbral) la célula responde activamente abriendo voltage-gated ion channels, que producen una respuesta todo o nada: el Potencial de Acción. (ver KSJ-Box 7.1, pag 127)
- (detrás: esta la parte derecha de la fig KSJsp-F2.8 (pag 32) )