
Recomendados
Recomendados
Más contenido relacionado
Similar a 7. GENERACION POTENCIALES BIOELECTRICOS.pdf
Similar a 7. GENERACION POTENCIALES BIOELECTRICOS.pdf (20)
Más de jorgequintero92
Más de jorgequintero92 (7)
Último
Último (20)
7. GENERACION POTENCIALES BIOELECTRICOS.pdf
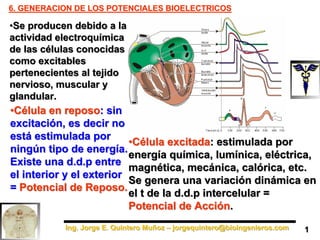
- 1. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6. GENERACION DE LOS POTENCIALES BIOELECTRICOS 6. GENERACION DE LOS POTENCIALES BIOELECTRICOS •Se producen debido a la actividad electroquímica de las células conocidas como excitables pertenecientes al tejido nervioso, muscular y glandular. •Se producen debido a la actividad electroquímica de las células conocidas como excitables pertenecientes al tejido nervioso, muscular y glandular. •Célula en reposo: sin excitación, es decir no está estimulada por ningún tipo de energía. Existe una d.d.p entre el interior y el exterior = Potencial de Reposo. •Célula en reposo: sin excitación, es decir no está estimulada por ningún tipo de energía. Existe una d.d.p entre el interior y el exterior = Potencial de Reposo. •Célula excitada: estimulada por energía química, lumínica, eléctrica, magnética, mecánica, calórica, etc. Se genera una variación dinámica en el t de la d.d.p intercelular = Potencial de Acción. •Célula excitada: estimulada por energía química, lumínica, eléctrica, magnética, mecánica, calórica, etc. Se genera una variación dinámica en el t de la d.d.p intercelular = Potencial de Acción. 1 1
- 2. 6.1 POTENCIAL DE REPOSO 6.1 POTENCIAL DE REPOSO - - MEMBRANA SEMIPERMEABLE MEMBRANA SEMIPERMEABLE Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Toda célula, animal o vegetal, se caracteriza por tener una membrana semipermeable y selectiva que demarca el límite de separación entre su estructura interna y el medio que la rodea (siempre líquido). Toda célula, animal o vegetal, se caracteriza por tener una membrana semipermeable y selectiva que demarca el límite de separación entre su estructura interna y el medio que la rodea (siempre líquido). Tanto el interior de la célula, como el medio ambiente que la rodea, son soluciones electrolíticas y el sistema se caracteriza por el hecho de que en el interior de la célula se acumula potasio, mientras en el medio externo, el ion prevaleciente es el sodio. Tanto el interior de la célula, como el medio ambiente que la rodea, son soluciones electrolíticas y el sistema se caracteriza por el hecho de que en el interior de la célula se acumula potasio, mientras en el medio externo, el ion prevaleciente es el sodio. ION CONCENTRACION CONCENTRACION INTERNA (m m ol/L) EXTERNA (m m ol/L) Na+ 12 145 K+ 155 4 Cl- 4 120 A- 155 A :A niones orgánicos diversos ION CONCENTRACION CONCENTRACION INTERNA (m m ol/L) EXTERNA (m m ol/L) Na+ 12 145 K+ 155 4 Cl- 4 120 A- 155 A :A niones orgánicos diversos 2 2
- 3. 6.2 POTENCIAL DE REPOSO 6.2 POTENCIAL DE REPOSO - - ASIMETRIA IONICA ASIMETRIA IONICA • Si estas concentraciones se igualaran, representaría la muerte de la célula. (A) • Ahora bien, esta asimetría iónica da origen a un potencial bioeléctrico, el denominado “Potencial de Membrana” o de reposo. • En general, cuando dos soluciones electrolíticas se encuentran separadas por una barrera semipermeable (membrana, película, restricción, etc.), se establece un potencial electroquímico. (B) • Si estas concentraciones se igualaran, representaría la muerte de la célula. (A) • Ahora bien, esta asimetría iónica da origen a un potencial bioeléctrico, el denominado “Potencial de Membrana” o de reposo. • En general, cuando dos soluciones electrolíticas se encuentran separadas por una barrera semipermeable (membrana, película, restricción, etc.), se establece un potencial electroquímico. (B) Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 3 3
- 4. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.3 POTENCIAL DE REPOSO 6.3 POTENCIAL DE REPOSO - - PARAMETROS DE PERMEABILIDAD PARAMETROS DE PERMEABILIDAD • La membrana celular es semipermeable: permite el flujo de algunos iones mientras impide el paso de otros. Es selectiva. • La membrana celular es semipermeable: permite el flujo de algunos iones mientras impide el paso de otros. Es selectiva. • Velocidades de los iones permeables: •Na+: 2 E- 8 cm/s (- permeable) •K+ : 2 E - 6 cm/s •Cl- : 4 E - 6 cm/s (+permeable) •Resto sustancias: 0 cm/s • Velocidades de los iones permeables: •Na+: 2 E- 8 cm/s (- permeable) •K+ : 2 E - 6 cm/s •Cl- : 4 E - 6 cm/s (+permeable) •Resto sustancias: 0 cm/s • En el estudio del potencial eléctrico de una célula solo se toman en cuenta los iones con mayor permeabilidad: Na+, K+, Cl-. • En el estudio del potencial eléctrico de una célula solo se toman en cuenta los iones con mayor permeabilidad: Na+, K+, Cl-. 4 4
- 5. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.4 POTENCIAL DE REPOSO 6.4 POTENCIAL DE REPOSO - - FACTORES QUE INTERVIENEN FACTORES QUE INTERVIENEN •Factores que intervienen en la generación del potencial de reposo: •Los gradientes de difusión •El CE opuesto a la difusión •La membrana semipermeable. •Factores que intervienen en la generación del potencial de reposo: •Los gradientes de difusión •El CE opuesto a la difusión •La membrana semipermeable. • Como {K+}i > {K+}o se presenta un gradiente de difusión, es decir una fuerza que tiende a que la {K+}i = {K+}o y por lo tanto hace que K+ salgan al exterior. • Como {K+}i > {K+}o se presenta un gradiente de difusión, es decir una fuerza que tiende a que la {K+}i = {K+}o y por lo tanto hace que K+ salgan al exterior. •El mismo fenómeno hará que iones Cl- ingresen a la célula mientras que los de Na+ por su baja permeabilidad en reposo prácticamente no se mueven. •El mismo fenómeno hará que iones Cl- ingresen a la célula mientras que los de Na+ por su baja permeabilidad en reposo prácticamente no se mueven. • En vista de lo anterior, el interior de la célula es mas negativo que el exterior, estableciéndose un CE dirigido hacia el interior que tiende a inhibir el flujo de K+ y Cl-. Las fuerzas de difusión y eléctricas se oponen entre si llegandose a un estado de equilibrio. • En vista de lo anterior, el interior de la célula es mas negativo que el exterior, estableciéndose un CE dirigido hacia el interior que tiende a inhibir el flujo de K+ y Cl-. Las fuerzas de difusión y eléctricas se oponen entre si llegandose a un estado de equilibrio. •El rango del potencial de reposo para varios tipos de células va desde -60mV a 100 mV. Una célula en estado de reposo se dice que está POLARIZADA. •El rango del potencial de reposo para varios tipos de células va desde -60mV a 100 mV. Una célula en estado de reposo se dice que está POLARIZADA. 5 5
- 6. 6.5 POTENCIAL DE REPOSO 6.5 POTENCIAL DE REPOSO - - GRADIENTE DIFUSIONS GRADIENTE DIFUSIONS Vs Vs CAMPO E CAMPO E Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6 6
- 7. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.6 POTENCIAL DE REPOSO 6.6 POTENCIAL DE REPOSO - - PARTICIPACION POR ION PARTICIPACION POR ION •Ecuación de NERNST del potencial de reposo: •Ecuación de NERNST del potencial de reposo: •Vk = equilibrium voltage for the kth ion across the membrane •R = gas constant [8.314 J/(mol·K)] •T = absolute temperature [K] • zk = valence of the kth ion •F = Faraday's constant [9.649 E04 C/mol] •ci,k = intracellular concentration of the kth ion •co,k = extracellular concentration of the kth ion •Vk = equilibrium voltage for the kth ion across the membrane •R = gas constant [8.314 J/(mol·K)] •T = absolute temperature [K] • zk = valence of the kth ion •F = Faraday's constant [9.649 E04 C/mol] •ci,k = intracellular concentration of the kth ion •co,k = extracellular concentration of the kth ion 7 7
- 8. 6.7 POTENCIAL DE REPOSO 6.7 POTENCIAL DE REPOSO - - PARTICIPACION TOTAL IONICA PARTICIPACION TOTAL IONICA •El equilibrio de Donnan: •El equilibrio de Donnan: •Ecuación de Goldman-Hodgkin-Katz del potencial de reposo: •Ecuación de Goldman-Hodgkin-Katz del potencial de reposo: Pk, Pna, Pcl: Permeabilidades Pk, Pna, Pcl: Permeabilidades Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 8 8
- 9. 6.8 POTENCIAL DE REPOSO 6.8 POTENCIAL DE REPOSO - - EJERCICIO EJERCICIO Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 9 9
- 10. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.9 POTENCIAL DE ACCION 6.9 POTENCIAL DE ACCION - - MANERA DE GENERARLO MANERA DE GENERARLO • La membrana celular se puede excitar por: • Corrientes iónicas que la atraviesen. •Algún tipo de energía aplicada externamente. • La membrana celular se puede excitar por: • Corrientes iónicas que la atraviesen. •Algún tipo de energía aplicada externamente. •Efecto en la membrana: cambia su permeabilidad, empezando a ser más permeable al Na+ •El potencial de membrana empieza a variar, dejando de ser estático (potencial de reposo), convirtiéndose en dinámico, generándose el potencial de acción. •Efecto en la membrana: cambia su permeabilidad, empezando a ser más permeable al Na+ •El potencial de membrana empieza a variar, dejando de ser estático (potencial de reposo), convirtiéndose en dinámico, generándose el potencial de acción. 10 10
- 11. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.10 POTENCIAL DE ACCION 6.10 POTENCIAL DE ACCION - - INTENSIDAD INTENSIDAD Vs Vs TIEMPO DEL ESTIMULO TIEMPO DEL ESTIMULO 11 11
- 12. 6.11 POTENCIAL DE ACCION 6.11 POTENCIAL DE ACCION - - COMPORTAMIENTO EN EL TIEMPO COMPORTAMIENTO EN EL TIEMPO Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 12 12
- 13. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.12 POTENCIAL DE ACCION 6.12 POTENCIAL DE ACCION - - DESPOLARIZACION DESPOLARIZACION • Proceso en la generación del potencial de acción: •1. Se aplica un estímulo a la membrana celular. • Proceso en la generación del potencial de acción: •1. Se aplica un estímulo a la membrana celular. 2. La membrana se hace muy permeable al Na+, presentandose una avalancha desde el exterior al interior, intentando alcanzar el equilibrio de { }. 2. La membrana se hace muy permeable al Na+, presentandose una avalancha desde el exterior al interior, intentando alcanzar el equilibrio de { }. 3. Al mismo tiempo los iones de K+ salen de la célula, a menor velocidad. El interior llega a ser más ligeramente mas positivo en el interior (+20mV). 3. Al mismo tiempo los iones de K+ salen de la célula, a menor velocidad. El interior llega a ser más ligeramente mas positivo en el interior (+20mV). Este proceso desde -80mV en reposo hasta +20 mV se llama DESPOLARIZACION. Este proceso desde -80mV en reposo hasta +20 mV se llama DESPOLARIZACION. 13 13
- 14. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.13 POTENCIAL DE ACCION 6.13 POTENCIAL DE ACCION - - REPOLARIZACION REPOLARIZACION Este proceso desde +20mV hasta -80 mV se llama REPOLARIZACION. Este proceso desde +20mV hasta -80 mV se llama REPOLARIZACION. •Proceso en la generación del potencial de acción: •4. La avalancaha de Na+ termina cuando las concentraciones han alcanzado el equilibrio. •Proceso en la generación del potencial de acción: •4. La avalancaha de Na+ termina cuando las concentraciones han alcanzado el equilibrio. 5. La membrana vuelve a ser impermeable al Na+ bloqueando el flujo de este. 5. La membrana vuelve a ser impermeable al Na+ bloqueando el flujo de este. •6. Mediante un proceso activo denominado la BOMBA Na-K, los iones Na+ son expulsados rápidamente al exterior y los K+ introducidos hasta que la membrana vuelve a su estado de reposo. (-80mV). •6. Mediante un proceso activo denominado la BOMBA Na-K, los iones Na+ son expulsados rápidamente al exterior y los K+ introducidos hasta que la membrana vuelve a su estado de reposo. (-80mV). 14 14
- 15. 6.14 POTENCIAL DE ACCION 6.14 POTENCIAL DE ACCION - - CANALES IONICOS CANALES IONICOS - - BOMBA BOMBA Na+K Na+K Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 15 15
- 16. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.15 POTENCIAL DE ACCION 6.15 POTENCIAL DE ACCION - - CARACTERISTICAS CARACTERISTICAS •En las células nerviosas y musculares el tiempo de repolarización (Trep) es muy pequeño y el tiempo del potencial de acción (Tpa) es aproximadamente de 1 ms. •En las células nerviosas y musculares el tiempo de repolarización (Trep) es muy pequeño y el tiempo del potencial de acción (Tpa) es aproximadamente de 1 ms. • En el músculo cardiaco el Trep está entre 150 y 300 ms. • En el músculo cardiaco el Trep está entre 150 y 300 ms. •El potencial de acción posee una amplitud y duración fijos, independientes de la intensidad y duración del estímulo ( siempre que este supere el umbral de excitación). LEY DEL TODO O NADA. •El potencial de acción posee una amplitud y duración fijos, independientes de la intensidad y duración del estímulo ( siempre que este supere el umbral de excitación). LEY DEL TODO O NADA. Tpa Tpa Trep Trep 16 16
- 17. 6.15 POTENCIAL DE ACCION 6.15 POTENCIAL DE ACCION - - CARACTERISTICAS CARACTERISTICAS •Tiempo refractario absoluto (Tra): Periodo de tiempo en el que es imposible volver a estimular la célula por mas que se aumente el valor de la intensidad del estímulo (1 ms en células nerviosas) •Tiempo refractario absoluto (Tra): Periodo de tiempo en el que es imposible volver a estimular la célula por mas que se aumente el valor de la intensidad del estímulo (1 ms en células nerviosas) •Tiempo refractario relativo (Trr): En este periodo se puede producir otro potencial de acción, pero se requiere aplicar un estímulo mayor que el inicial. •Tiempo refractario relativo (Trr): En este periodo se puede producir otro potencial de acción, pero se requiere aplicar un estímulo mayor que el inicial. Tra Tra Trr Trr Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 17 17
- 18. 6.16 POTENCIAL DE ACCION 6.16 POTENCIAL DE ACCION - - PROPAGACION PROPAGACION •La despolarización empieza en una parte determinada de la membrana y las corrientes iónicas que circulan al generarse un potencial de acción excitan áreas adyacentes de la célula, propagandose la excitación (despolarización). Esta excitación puede propagarse a otras células vecinas. •La despolarización empieza en una parte determinada de la membrana y las corrientes iónicas que circulan al generarse un potencial de acción excitan áreas adyacentes de la célula, propagandose la excitación (despolarización). Esta excitación puede propagarse a otras células vecinas. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com •La velocidad a la que se mueve un potencial de acción se denomina velocidad de propagación. En los nervios (20 a 140 m/s) y en el músculo cardiaco (0.2 a 0.4 m/s). •La velocidad a la que se mueve un potencial de acción se denomina velocidad de propagación. En los nervios (20 a 140 m/s) y en el músculo cardiaco (0.2 a 0.4 m/s). 18 18
- 19. 6.17 INTERCONEXION DE MOTONEURONAS CON FIBRAS 6.17 INTERCONEXION DE MOTONEURONAS CON FIBRAS MUSCULARES ( MUSCULARES (English English) ) Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 19 19
- 20. 6.18 .INTERCONEXION DE MOTONEURONAS CON FIBRAS 6.18 .INTERCONEXION DE MOTONEURONAS CON FIBRAS MUSCULARES ( MUSCULARES (Spanish Spanish) ) Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 20 20
- 21. Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com Ing. Jorge E. Quintero Muñoz – jorgequintero@bioingenieros.com 6.19 POTENCIALES BIOELECTRICOS 6.19 POTENCIALES BIOELECTRICOS - - CONCLUSION CONCLUSION •Aunque la medida de un PA es posible, se trata de una medida difícil pues requiere la colocación de un electrodo dentro de una célula. •Aunque la medida de un PA es posible, se trata de una medida difícil pues requiere la colocación de un electrodo dentro de una célula. •En diagnóstico clínico se miden y registran los efectos combinados de un número elevado de PA empleando electrodos superficiales o de aguja insertados en un músculo, nervio o alguna parte del cerebro. •En diagnóstico clínico se miden y registran los efectos combinados de un número elevado de PA empleando electrodos superficiales o de aguja insertados en un músculo, nervio o alguna parte del cerebro. •La medida de un potencial bioeléctrico cualquiera es el resultado de la sumatoria en el tiempo de los PA externos de un elevado número de células: Cardiacas (ECG), Musculares (EMG), Cerebrales (EEG), etc. •La medida de un potencial bioeléctrico cualquiera es el resultado de la sumatoria en el tiempo de los PA externos de un elevado número de células: Cardiacas (ECG), Musculares (EMG), Cerebrales (EEG), etc. 21 21