Descargar para leer sin conexión























































































![LA REPRESENTACIÓN NEUROLÓGICA
José Elías Delgado B. XIV – Master C.O.I 89
CORTEZA VISUAL EXTRAESTRIADA.
La corteza estriada realiza un análisis básico de las aferencias geniculadas y
transmite, canales de codificación del mensaje visual (codificaciones criticas)a
zonas corticales periestriadas superiores para su ulterior interpretación.
En el área V2 se analiza de forma global, la información procedente de V1,
independizándola total o parcialmente de la variable tiempo, haciendo posible
nuevas codificaciones que permiten una traducción estable invariable de los
objetos reales. La codificación receptiva se hace mas especifica en la
discriminación de movimiento, posición, constancia de tamaño, claridad, color y
textura. Mal codificadas por las repetidas modificaciones de la imagen retenida
en la vía visual.
En la representación visual de códigos de procesamiento de información, la
primera representación simbólica completa de la imagen se desencadena entre
las conexiones estriadas V1 y periestriada V2 las cuales contienen un conjunto
de casillas en donde se analiza por secciones el campo de visión completo, aquí
se integran y se recomponen las diferentes señales antes de remitirlas a áreas
visuales superiores, (difirenciándolas de los distintos atributos de la imagen y
propios de la visión).
Hace algunos años el conocimiento de la fisiología postulada que la percepción
tenia lugar en áreas visuales de asociaciones que rodean la corteza estriada y que
Brodmann denomina áreas 18 y 19.
Los estudios histológicos, electrofisiológicos y de alta tecnología con aplicación
de marcadores radio activos [TOMOGRAFIA POR EMISION DE
POSITRONES - TEP] En otros, revelan que las áreas 18 y 19 contienen al
menos 5 áreas corticales distintas dedicadas al procesamiento visual V2 – VSA
– V4 y V5. Siguen sin localizarse ni caracterizarse obras áreas visuales en, y mas
allá de las áreas 18 y 19.](https://image.slidesharecdn.com/fundamentosneuronales1-150203122939-conversion-gate02/85/Fundamentos-neuronales-1-89-320.jpg)



![LA REPRESENTACIÓN NEUROLÓGICA
José Elías Delgado B. XIV – Master C.O.I 93
ÁREA EXTRAESTRIADA V5
Las células de V5 o área temporal media TM, presentan una citoarquitectura
definida por el [] elevado de Mielina.
ALLMAN Y KASS.
La obtención de registros de esta zona representa el hemicampo de visión
contralateral.
El área V5 de la corteza extraestriada contiene neuronas exquisitamente
sensibles al movimiento del estimulo.
Área V5 <<área codificadora del movimiento>>
La unidad de área V5recibe aferencias desde la capa IVB de V1 y desde la
gruesa capa citocromooxidasa de V2. Estas dos regiones aferentes están
pobladas por unidades selectivas a la dirección.
Ø El área V5 <<codificadora del movimiento>> de la corteza extraestriada
controla los mecanismos de seguimiento suave y la percepción del
movimiento visual.](https://image.slidesharecdn.com/fundamentosneuronales1-150203122939-conversion-gate02/85/Fundamentos-neuronales-1-93-320.jpg)








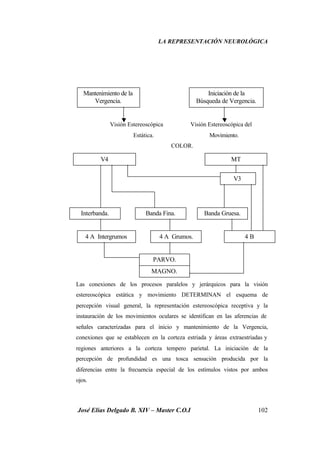




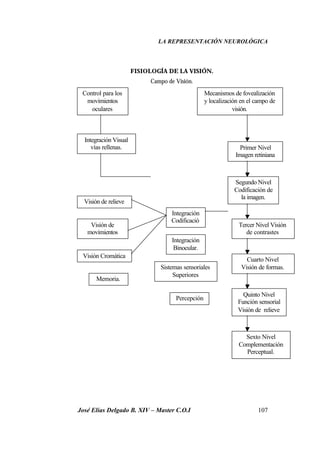




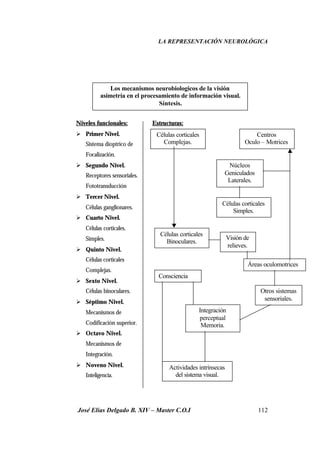


















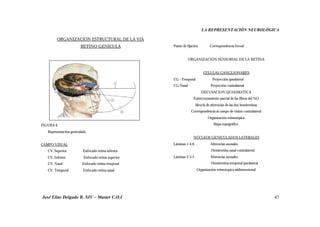
Este documento resume investigaciones sobre la representación neurológica de la función visual. Detalla los hallazgos de pioneros como Broca y Lashley sobre la lateralización cerebral y las diferencias funcionales entre hemisferios. También revisa estudios más recientes sobre la organización cerebral de la percepción visual realizados por Hubel, Wiesel, Marr y otros. El objetivo es establecer un marco teórico sobre la actividad neuronal involucrada en la visión y su relación con la optometría clínica.











































































