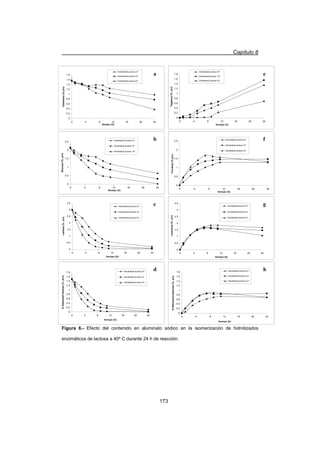
Este capítulo describe el proceso de hidrólisis enzimática de lactosa en leche y permeados de lactosuero utilizando β-galactosidasa de Bacillus circulans inmovilizada en resinas acrílicas entrampadas en una matriz de alginato. La hidrólisis enzimática de lactosa es una alternativa al método químico para aprovechar la lactosa en la producción de jarabes de glucosa y galactosa, los cuales tienen mayor poder edulcorante y solubilidad que la lactosa. El proceso enzimático puede realizarse a temper


























































![Capítulo 3
57
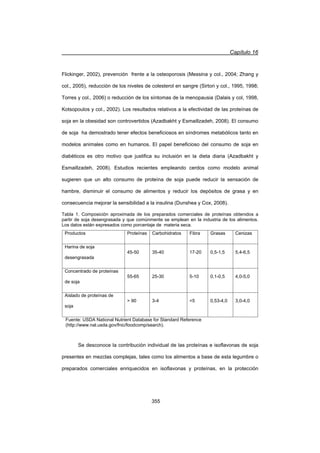
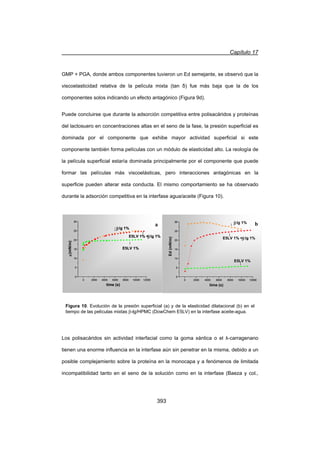
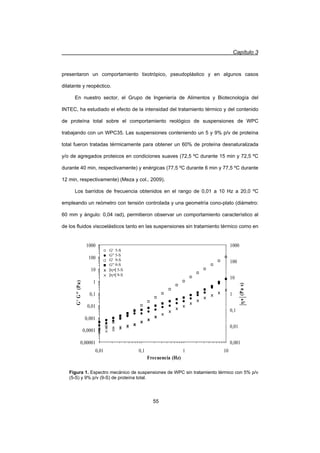
A frecuencias superiores a la frecuencia de corte (Ȧc), se visualizó un comportamiento
predominantemente sólido dónde G' fue mayor a G. Además, en las suspensiones se
observó una independencia de G' y una escasa dependencia de G con el contenido
de proteína total en el rango de frecuencias estudiado (Figuras 1 y 2).
Tabla 1. Tiempos característicos de relajación de suspensiones de WPC obtenidos a partir del
modelo de Ley de Potencia.
Muestras
Tiempos característicos de
ralajación [s]*
5-S1
7,25±0,73a
9-S1
3,51±0,02bc
5-72,52
3,68±0,29b
9-72,53
0,64±0,04d
5-77,52
2,43±0,08e
9-77,53
0,63±0,05d
* Letras diferentes en cada columna indica
diferencias significativas (p0,05).
1
Suspensiones de WPC sin tratamiento con 5% p/v
(5-S) y 9% p/v (9-S) de proteína total.
2
Suspensiones de WPC con 5% p/v de proteína total
tratadas térmicamente en condiciones suaves (5-
72,5) y en condiciones enérgicas (5-77,5).
3
Suspensiones de WPC con 9% p/v de proteína total
tratadas térmicamente en condiciones suaves (9-
72,5) y en condiciones enérgicas (9-77,5).
La poca dependencia de los módulos con la concentración también ha sido
observada en algunas dispersiones coloidales de proteínas globulares en estado
nativo y fue interpretada en términos de la formación de coloides cristalinos (Ikeda y
Nishinari, 2001a, b; Ikeda y col., 2001). Los espectros mecánicos de las suspensiones
tratadas térmicamente para un mismo contenido de proteína total presentaron una
descripción similar al de las suspensiones sin tratamiento térmico. No obstante, existió
una dependencia de G con la intensidad del tratamiento térmico que se analizó
empleando los tiempos característicos de relajación (tc) calculados como la inversa de
Ȧc (Tabla 1). Además, en todas las suspensiones de WPC se observó un
comportamiento dilatante, dónde la viscosidad compleja (|K*|) se incrementó con la
frecuencia (Figura 1).](https://image.slidesharecdn.com/funsionalidaddeproteinasdeleche-141105103942-conversion-gate01/85/Funsionalidad-de-proteinas-de-leche-63-320.jpg)

![Capítulo 3
59
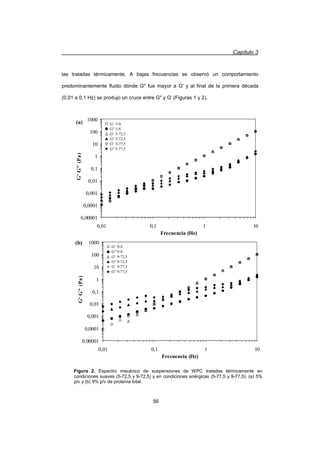
partículas producida por la presencia de fuerzas hidrodinámicas elevadas provocan la
presencia de un comportamiento dilatante.
Los parámetros reológicos derivados del modelo de Ley de Potencia para G' y
G (G'=aȦx
, G=bȦy
) permiten analizar el comportamientos de las suspensiones y
comparar tratamientos. Los coeficientes a y b representan las magnitudes de G' y G
respectivamente a una dada frecuencia (1 rad/s) y los exponentes x e y representan la
velocidad de variación de los módulos con la frecuencia. Por ejemplo, un exponente x
cercano a cero indica que G' no varía con la frecuencia, representando el
comportamiento de un sólido viscoelástico, como por ejemplo un gel (Resch y Daubert,
2002). La matriz de un gel tiene la característica de poseer un alto grado de
estructuración, por este motivo ejerce una alta resistencia a la deformación que se
pone en evidencia con una menor velocidad de variación de los módulos con la
frecuencia.
En el caso de las suspensiones analizadas, el exponente x fue significativamente
mayor que y en todas las suspensiones analizadas, indicando que G' se incrementa
más rápidamente que G al aumentar la frecuencia (Tabla 2).
Tabla 2. Parámetros reológicos de las suspensiones de WPC obtenidos a partir del modelo de
Ley de Potencia.
Muestras
Módulo elástico
(G')
Módulo viscoso
(G)
a [Pa s]* x [-]* R2
b [Pa s]* y [-]* R2
5-S1
0,025±0,000a
2,011±0,007a
0,99 0,005±0,000a
1,177±0,016ab
0,98
9-S1
0,024±0,000a
2,019±0,007a
0,99 0,008±0,000a
1,119±0,008c
0,99
5-72,52
0,025±0,001a
2,005±0,009a
0,99 0,008±0,002a
1,106±0,007c
0,99
9-72,53
0,019±0,000a
2,114±0,008a
0,99 0,033±0,002b
0,872±0,021e
0,99
5-77,52
0,026±0,000a
1,987±0,003a
0,99 0,011±0,000a
1,054±0,006d
0,99
9-77,53
0,041±0,009b
1,786±0,093b
0,99 0,066±0,009c
0,728±0,039f
0,96
* Letras diferentes en cada columna indica diferencias significativas (p0,05).
1
Suspensiones de WPC sin tratamiento con 5% p/v (5-S) y 9% p/v (9-S) de proteína total.
2
Suspensiones de WPC con 5% p/v de proteína total tratadas térmicamente en condiciones suaves (5-
72,5) y en condiciones enérgicas (5-77,5).
3
Suspensiones de WPC con 9% p/v de proteína total tratadas térmicamente en condiciones suaves (9-
72,5) y en condiciones enérgicas (9-77,5).](https://image.slidesharecdn.com/funsionalidaddeproteinasdeleche-141105103942-conversion-gate01/85/Funsionalidad-de-proteinas-de-leche-65-320.jpg)





![Capítulo 3
65
maduración.
Los parámetros reológicos derivados del modelo de Ley de Potencia para G' y
G permiten analizar el comportamiento reológico durante la maduración de los
quesos. El exponente x fue significativamente mayor que y en todas las muestras de
queso analizadas, indicando que G' se incrementa más rápidamente que G al
aumentar la frecuencia (Tabla 3).
Tabla 3. Parámetros reológicos obtenidos a partir del modelo de Ley de Potencia para el
módulo elástico (G'=aZx
) y el módulo viscoso (G=bZy
) de un queso blando elaborado con
proteínas del suero.
Tiempo de
maduración
[días]
Módulo elástico
(G')
Módulo viscoso
(G)
a [Pa s]* x [-]* R2
b [Pa s]* y [-]* R2
1 121,1±5,2a
0,217±0,001a
0,99 39,5±1,5a
0,185±0,002a
0,99
21 79,2±16,1b
0,231±0,011ab
0,99 28,8±5,5b
0,210±0,011b
0,99
48 72,1±10,6b
0,236±0,008ab
0,99 25,6±3,5bc
0,233±0,008b
0,99
76 43,9±4,5c
0,248±0,017b
0,99 17,7±1,5c
0,215±0,011b
0,99
*Letras diferentes en cada columna indica diferencias significativas (P0,05).
Durante la maduración de los quesos, se observó una tendencia decreciente en
los parámetros a y b, encontrándose diferencias significativas entre los valores de a de
los días 1 y 48 de maduración y entre los valores de b de los días 21 y 76. Por otro
lado, también se observó que los valores de ambos exponentes x e y aumentaron
significativamente entre los días 1 y 21 de maduración, permaneciendo constantes el
resto del tiempo. Estas observaciones demostraron que los tanto G' como G
presentaron una mayor velocidad de variación con la frecuencia durante la
maduración, indicando que disminuyó el grado de estructuración de la red de caseína
(Gravier y col., 2004). Además, los valores de los módulos G' y G a una frecuencia
determinada (representado por los valores de a y b) disminuyeron durante la
maduración. Los productos provenientes de la hidrólisis de las caseínas son solubles
en agua y no contribuyen a formar la red de caseína (Ustunol y col., 1995). De esta
forma, la pérdida de estructura de las proteínas debido a la proteólisis implica una](https://image.slidesharecdn.com/funsionalidaddeproteinasdeleche-141105103942-conversion-gate01/85/Funsionalidad-de-proteinas-de-leche-71-320.jpg)














































































































































































































![Capítulo 13
281
h a 37ºC. El espectro de acción del fragmento f(63-117) sintetizado químicamente
incluye la inhibición de bacterias Gram-positivas, Gram-negativas y levaduras. Estos
resultados podrían ser relevantes desde el punto de vista fisiológico ya que apoyarían
la hipótesis de que los péptidos antimicrobianos se generan a partir de la leche
humana durante la digestión del lactante y, de ese modo juegan un papel muy
importante en el sistema defensivo del recién nacido. La kappacina (Malkoski y col.,
2001) es otro ejemplo de péptido antibacteriano derivado de la N-caseína. La
kappacina es la forma no glicosilada y fosforilada del CMP [N-caseína (106-169)]. Para
caracterizar la región activa de la kappacina, el péptido se hidrolizó con
endoproteinasa Glu-C, observándose que el péptido Ser (P)149
N-caseína A f(138-158)
era la forma activa con actividad antimicrobiana frente a Streptococcus mutans,
Escherichia coli y Pseudomonas gingivalis. Es muy importante destacar que la forma
activa corresponde al péptido no-glicosilado y fosforilado, ya que está demostrado que
las formas glicosiladas y no fosforiladas no tienen actividad frente a Streptococcus
mutans. Los estudios de modelización molecular y predicciones de estructura
secundaria de la kappacina han indicado que el péptido dispone de residuos que
podrían formar una D-hélice anfipática, y agregarse para formar un poro aniónico, de
modo que se incremente la permeabilidad de la membrana a los cationes. Dashper y
col., (2005) han observado que a pH 6,5, la kappacina es capaz de aumentar la
permeabilidad de los liposomas, por lo que se propuso un modo de acción
membranolítico, de manera que el péptido se agregaría para formar un poro aniónico.
La fosforilación, que como se comentó previamente es esencial para la actividad,
parece que es la responsable de un cambio de conformación en el péptido a través de
repulsiones electroestáticas o bien por unión con iones divalentes que le permite
interactuar con la membrana bacteriana. El CMP se ha detectado en estómago,
duodeno y yeyuno de humanos tras la ingestión de leche (Ledoux y col., 1999). La](https://image.slidesharecdn.com/funsionalidaddeproteinasdeleche-141105103942-conversion-gate01/85/Funsionalidad-de-proteinas-de-leche-287-320.jpg)

![Capítulo 13
283
péptidos se obtuvieron a partir de la hidrólisis de lactoferrina con pepsina y
demostraron mayor actividad antimicrobiana que la proteína precursora. En el caso de
la lactoferrina humana el péptido correspondía al fragmento f(1-47) y se denominó
lactoferricina humana, mientras que en la lactoferrina bovina se identificó el fragmento
f(17-41) como el responsable de la actividad (conocido como lactoferricina bovina).
Además, las dos lactoferricinas no actuaban siguiendo el mecanismo de acción
descrito para la lactoferrina que se basa en su capacidad para secuestrar hierro.
La lactoferricina se ha revelado como un péptido antimicrobiano con un amplio
espectro de actividades frente a bacterias Gram-positivas y Gram-negativas (Bellamy y
col., 1992), hongos (Muñoz y Marcos, 2006) y parásitos (León-Sicario y col., 2006).
Además, también se presenta actividad antiviral (Pietrantoni y col., 2006), antitumoral
(Iigo y col., 1999) y antiinflamatoria (Levay y Viljoen, 1995).
La hidrólisis de lactoferrina con pepsina provoca no sólo la liberación de lactoferricina y
fragmentos que contienen lactoferricina, sino también de otros péptidos catiónicos.
Estos fragmentos corresponden a lactoferrina f(277-288), lactoferrina f(267-285) y
lactoferrina f(267-288) (Recio y Visser, 1999b). Un péptido, f(268-284), englobado en
uno de estos fragmentos de lactoferrina y conocido como lactoferrampina ha sido
sintetizado observándose que presentaba actividad frente a Candida así como frente a
Bacillus subtilis, Escherichia coli y Pseudomonas aeruginosa (van der Kraan y col.,
2004). Además de pepsina porcina, se ha comprobado que la hidrólisis con otras
enzimas como la quimosina da lugar a la formación de fragmentos análogos a la
lactoferricina (Hoek y col., 1997). Recientemente, Haney y col., (2009) sintetizaron
lactoferrampina humana [f(269-285)] además del mismo péptido con diferentes
aminoácidos en los extremos N- y C-terminal. Estos investigadores observaron que un
aumento de la carga neta positiva del extremo C-terminal del péptido incrementaba
significativamente su actividad antimicrobiana y su actividad frente a Candida albicans.
Además de la lactoferrina humana y bovina, se ha empleado lactoferrina de diversos](https://image.slidesharecdn.com/funsionalidaddeproteinasdeleche-141105103942-conversion-gate01/85/Funsionalidad-de-proteinas-de-leche-289-320.jpg)

































![Capítulo 14
317
Respecto a las proteínas de suero, la hidrólisis con una combinación de
enzimas digestivas u otras enzimas con alto poder proteolítico y baja especificidad,
como la termolisina o la proteinasa K, permite la producción de hidrolizados con un
gran número de potentes péptidos inhibidores de la ECA (Abubakar y col., 1998;
Hernández-Ledesma y col., 2002; Mullally y col., 1997; Nurminen y col., 2000;
Pihlanto-Leppälä y col., 2000). Por ejemplo, el péptido denominado E-lactosin A [IPA,
f(78-80)], se ha identificado en un hidrolizado de E-lactoglobulina con proteinasa K.
Aunque este péptido mostró una moderada actividad IECA, presentó una potente
actividad antihipertensiva en ratas SHR (Abubakar y col., 1998). En un estudio con un
hidrolizado de E-lactoglobulina de cabra preparado con termolisina, se identificaron
dos potentes inhibidores de la ECA, de secuencias LLF y LQKW, y posteriormente, se
demostró su efecto antihipertensivo en ratas SHR (Hernández-Ledesma y col., 2002 y
2007a).
La hidrólisis de D-lactalbúmina con enzimas gástricas y pancreáticas produjo la
liberación del tetra-péptido YGLF que presentó actividad opiácea y antihipertensiva en
ratas normotensas y SHR, siendo esta última actividad debida a su efecto como
antagonista de receptores opiáceos (Ijäs y col., 2004). Estos resultados sugieren que
otros mecanismos de acción, diferentes a la inhibición de la ECA pueden estar
involucrados en el efecto antihipertensivo de péptidos derivados de proteínas lácteas.](https://image.slidesharecdn.com/funsionalidaddeproteinasdeleche-141105103942-conversion-gate01/85/Funsionalidad-de-proteinas-de-leche-323-320.jpg)



![Capítulo 14
321
Gómez-Ruiz, J. A., Ramos, M., Recio, I. (2002). Angiotensin-converting enzyme-
inhibitory peptides in Manchego cheeses manufactured with different starter
cultures. International Dairy Journal, 12, 697-706.
Gómez-Ruiz, J. A., Ramos, M., Recio, I. (2004a). Angiotensin converting enzyme-
inhibitory activity of peptides isolated from Manchego cheese. Stability under
simulated gastrointestinal digestion. International Diary Journal 14, 1075-1080.
Gómez-Ruiz, J. A., Recio, I., Belloque, J. (2004b). ACE inhibitory activity and structural
properties of peptide Asp-Lys-Ile-His-Pro [E-CN f(47–51)]. Study of the peptide
forms synthesised by different methods. Journal of Agricultural and Food Chemistry,
52, 6315–6319.
Gómez-Ruiz, J. A, Taborda, G., Amigo, L., Recio, I., Ramos, M., (2006). Identification
of ACE-inhibitory peptides in different Spanish cheeses by tandem mass
spectrometry. European Food Research Technology, 223, 595-601.
Hartmann, R., Meisel, H (2007). Food-derived peptides with biological activity: from
research to food applications. Current opinion in Biotechnology, 18, 163-169.
Hata, Y., Yamamoto, N., Ohni, M., Nakajima, K., Nakamura, Y., Takano, T. (1996). A
placebo-controlled study of the effect of sour milk on blood pressure in hypertensive
subjects. American Journal of Clinical Nutrition, 64, 767-771.
Hernández-Ledesma, B., Amigo, L., Ramos, M., Recio, I. (2004). Angiotensin
converting enzyme inhibitory activity in commercial fermented products. Formation
of peptides under simulated gastrointestinal digestion. Journal of Agricultural and
Food Chemistry, 52, 1504-1510.
Hernández-Ledesma, B., López-Expósito. I, Ramos, M., Recio, I. (2006). Bioactive
peptides from milk proteins. In Immunochemistry in Dairy Research, 37-60.
Research Signpost ISBN:81-308-0102-7.
Hernández-Ledesma, B., Miguel, M., Amigo, L., Aleixandre, M. A., Recio, I., (2007a).
Effect of simulated gastrointestinal digestion on the anti-hypertensive properties of
synthetic E-lactoglobulin peptide sequences. Journal of Dairy Research, 74, 336-
339.
Hernández-Ledesma, B., Recio, I., Amigo, L. (2007b). E-lactoglobulin as source of
bioactive peptides. Amino Acids, 35, 257-265.
Hernández-Ledesma, B., Recio, I., Ramos, M., Amigo, L. (2002). Preparation of ovine
and caprine E-lactoglobulin hydrolysates with ACE-inhibitory activity. Identification of
active peptides from caprine E-lactoglobulin hydrolysed with thermolysin.
International Dairy Journal, 12, 805-812.](https://image.slidesharecdn.com/funsionalidaddeproteinasdeleche-141105103942-conversion-gate01/85/Funsionalidad-de-proteinas-de-leche-327-320.jpg)
























![Capítulo 15
348
2004. Antimicrobial potential of probiotic or potentially probiotic lactic acid bacteria,
the first results of the international European research project PROPATH of the
PROEUHEALTH cluster. Microbiology Ecology Health Disease 16: 125-130.
De Vuyst, L., Vanderveken, F., van de Ven, S., Degeest, B. 1998. Production by and
isolation of exopolysaccharides from Streptococcus thermophilus grown in a milk
medium and evidence for their growth-associated biosynthesis. Journal of Applied
Microbiology 84: 1059-1068.
De Wit, J. N. 1998. Nutritional and functional characteristics of whey proteins in food
products. Journal of Dairy Science 81: 597-602.
Degeest, B., De Vuyst, L. 1999. Indication that the nitrogen source influences both
amount and size of exopolysaccharides produced by Streptococcus thermophilus
LY03 and modelling of the bacterial growth and exopolysaccharide production in a
complex medium. Applied and Environmental Microbiology 65: 2863-2870.
Degeest, B., Vaningelge M., F., Laws, A., De Vuyst, L. 2001 UDP-N-acetylglucosamine
4-epimerase activity indicates the presence of N-acetylgalactosamine in
exopolysaccharides of Streptococcus thermophilus strains. Applied and
Environmental Microbiology 67: 3976–3984.
Dinakar, P., Mistry V. V. 1994. Growth and viability of Bifidobacterium bifidum in
Cheddar cheese. Journal of Dairy Science 77:2854-2864.
Duboc, P., Mollet, B. 2001. Applications of exopolysaccharides in the dairy industry.
International Dairy Journal 11: 759-768.
Eigel, W. N., Butler, J. E., Ernstrom, C. A., Farrell, H. M., Harwalkar, V. R., Jenness, R.
Whitney, R. M. 1984. Nomenclature of proteins of cow’s milk: Fifth revision. Journal
of Dairy Science 67: 1599-1631.
Evans, E. W. 1982. Uses of milk proteins in formulated foods. In B. J. F. Hudson,
Developments in Food Proteins. London: Applied Science.
FAO, 2001. Food and Agriculture Organization of the United Nations. World Health
Organization. Evaluation of health and nutritional properties of probiotics in food
including powder milk with live lactic acid bacteria: Report of a Joint FAO/WHO
Expert Consultation, Córdoba, Argentina, 2001 [cited 2005 Dec 15]. Available from:
ftp://ftp.fao.org/es/esn/food/probio_ report_en.pdf.
Furtado, M. M., Partridge, J. A., Ustunol, Z. 1993. Ripening characteristics of reduced
fat cheddar cheese using heat-shocked and live Lactobacillus casei as adjunt
culture. (Abstr) Journal of Dairy Science 76 (Sup.1):101.](https://image.slidesharecdn.com/funsionalidaddeproteinasdeleche-141105103942-conversion-gate01/85/Funsionalidad-de-proteinas-de-leche-354-320.jpg)