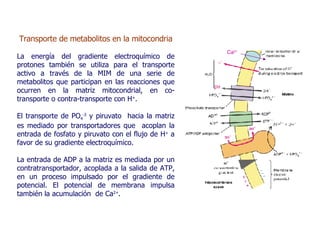
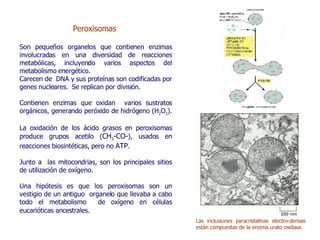
Las mitocondrias son organelos donde se producen reacciones de oxidación que generan la mayor parte del ATP en la célula. Contienen dos membranas que definen un espacio intermembrana y una matriz. En la matriz ocurren las reacciones del ciclo del ácido cítrico y la fosforilación oxidativa en la membrana interna, la cual usa un gradiente de protones para sintetizar ATP. Las mitocondrias pueden usar piruvato o ácidos grasos como combustible para generar acetil-CoA y almacenar energía











































![Transcripsion en eucariotas[1]](https://cdn.slidesharecdn.com/ss_thumbnails/transcripsioneneucariotas1-121021194324-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)




























































