Descargado 636 veces




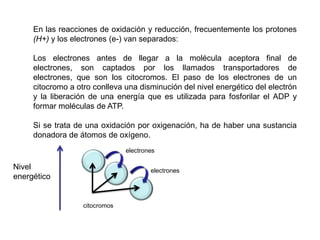









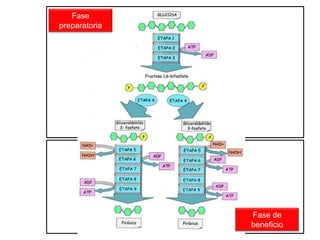







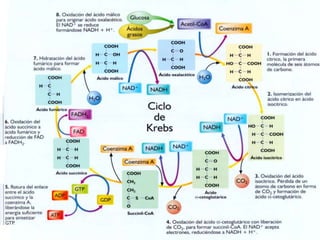


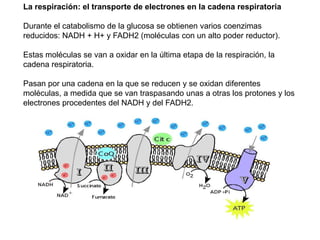





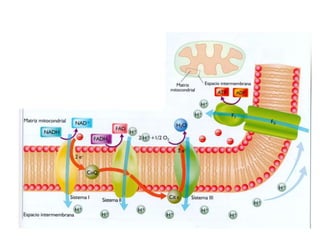
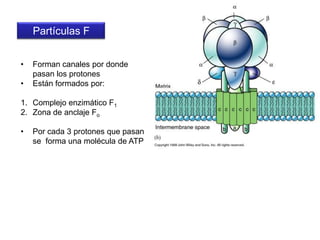




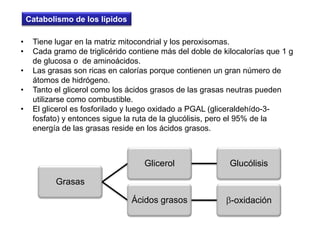

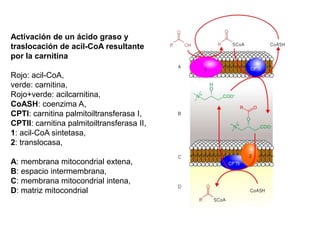


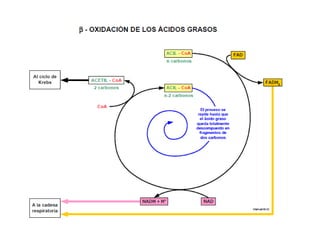

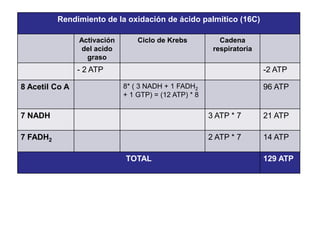



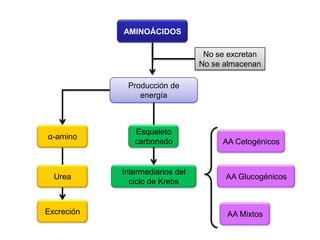



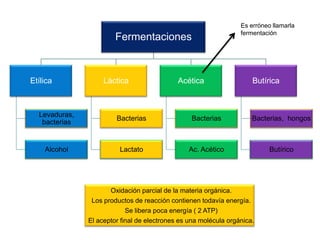










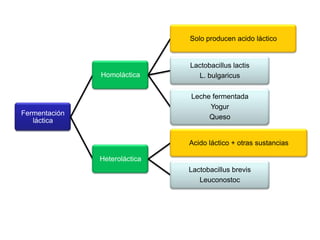



El documento describe los procesos de catabolismo aeróbico y anaeróbico. Explica que el catabolismo incluye la glucólisis, el ciclo de Krebs y la cadena respiratoria, donde se liberan electrones que se usan para producir ATP. También describe las fermentaciones anaeróbicas, donde un compuesto orgánico en lugar de oxígeno acepta los electrones, liberando menos energía.











































































































