Descargado 66 veces

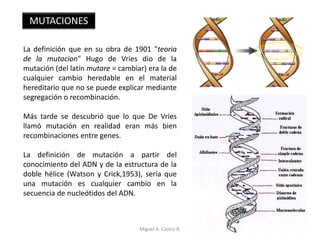
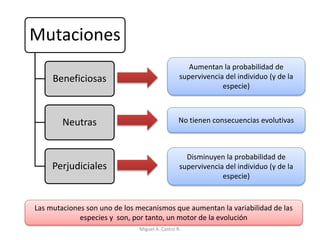
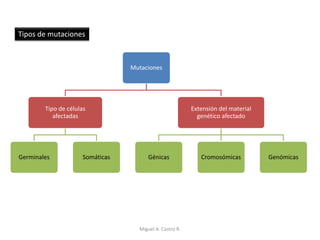
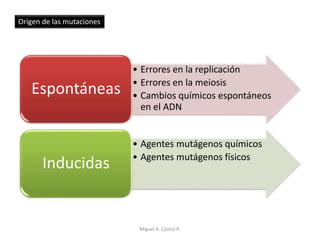

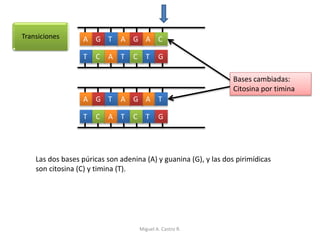

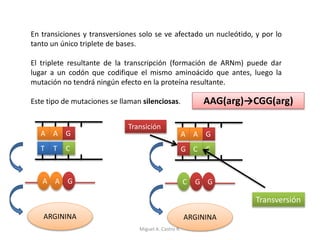
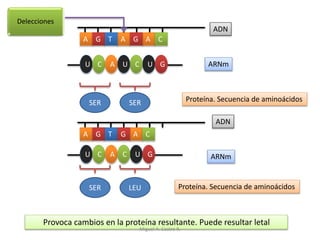
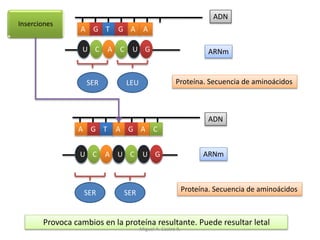
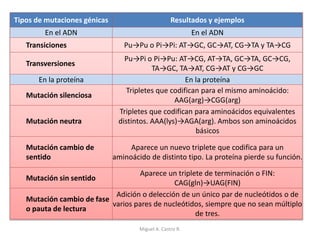
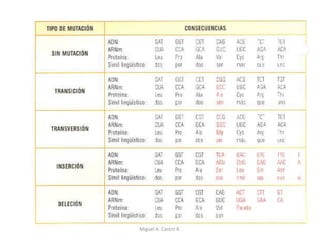

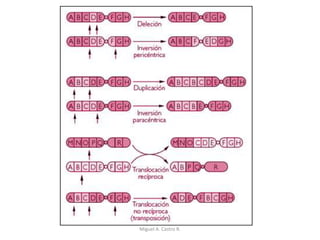
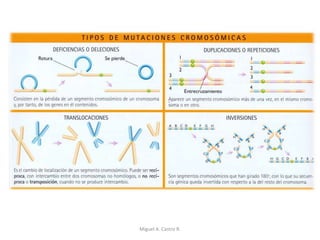

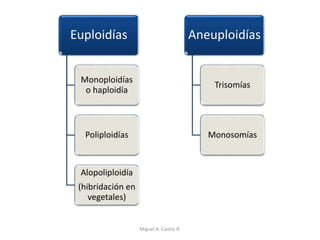
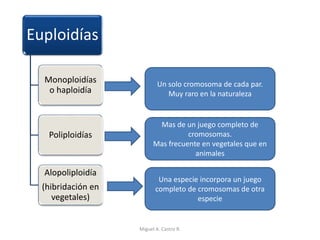
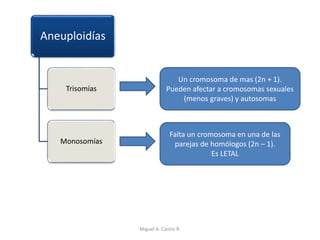
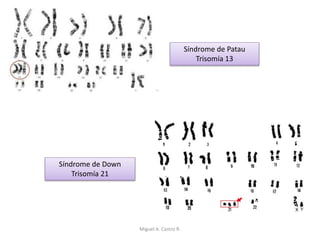

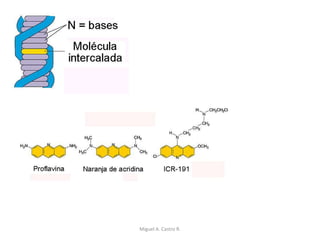
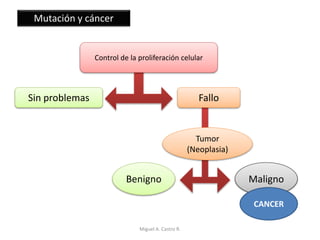

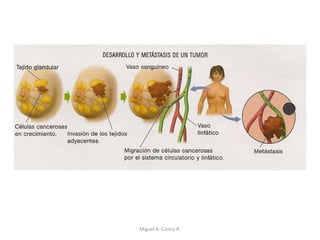
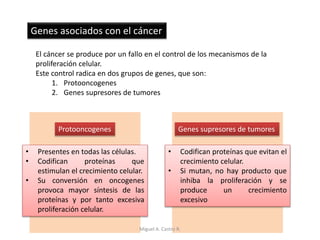
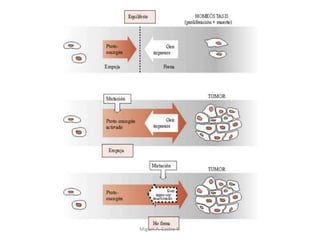

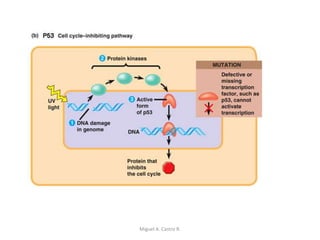

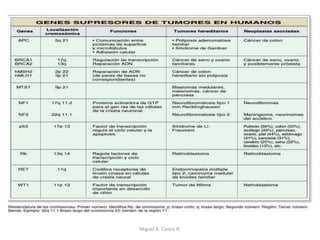

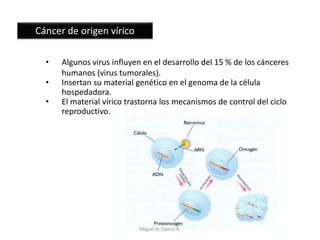
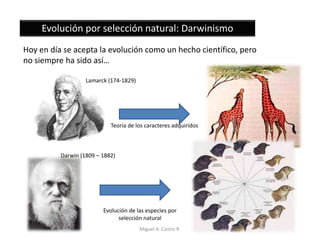


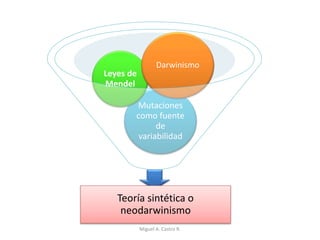



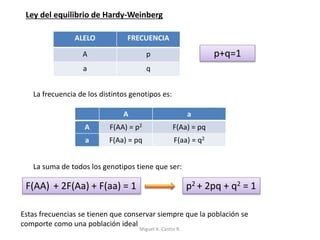

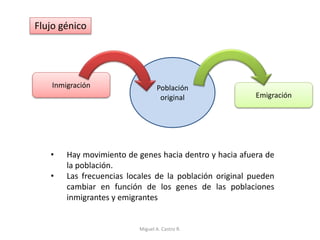


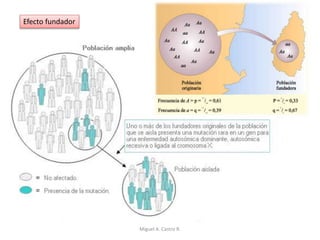



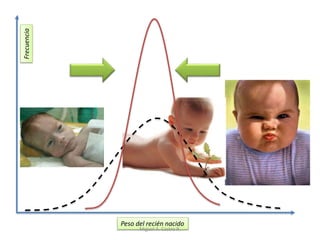
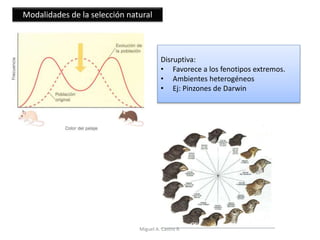
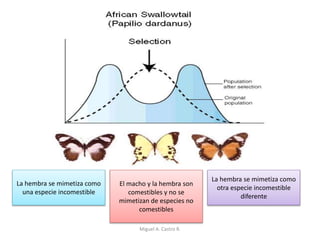

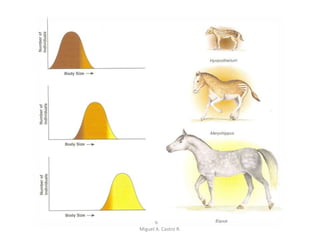

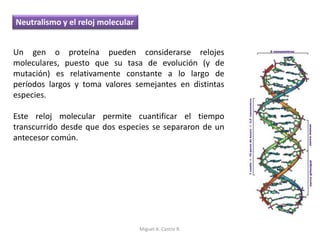

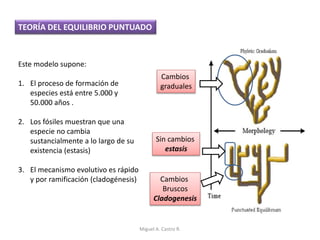





Este documento discute los conceptos de genética y evolución. Explica que las mutaciones son cambios hereditarios en el material genético que aumentan la variabilidad entre especies y son un motor de la evolución. Las mutaciones pueden ser beneficiosas, neutras o perjudiciales, y afectan a los genes o cromosomas. También discute cómo las mutaciones pueden causar cáncer al alterar los protooncogenes y genes supresores de tumores.























































































