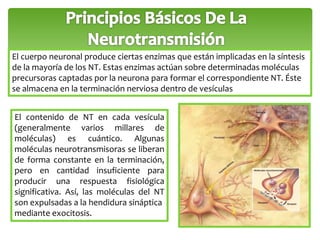
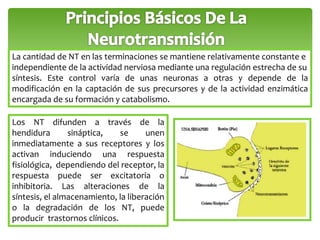
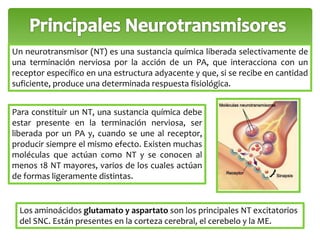
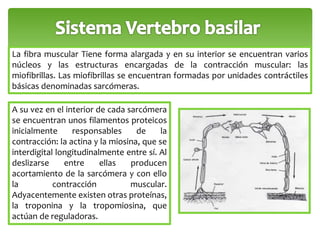
Este documento trata sobre los neurotransmisores y las células del sistema nervioso. Explica que las neuronas transmiten señales a través de redes de comunicación utilizando neurotransmisores como el glutamato, GABA, dopamina y serotonina. También describe los diferentes tipos de receptores de neurotransmisores y cómo regulan la liberación y recaptación de los neurotransmisores.