Bioquimica (unidad iv)
•Descargar como DOC, PDF•
3 recomendaciones•764 vistas

El documento describe el ciclo de la urea, que es el proceso mediante el cual el hígado convierte el amoníaco tóxico producido durante el metabolismo de proteínas en urea, la cual es menos tóxica y puede ser excretada por los riñones. El ciclo de la urea consta de cinco reacciones enzimáticas que tienen lugar en la mitocondria y el citosol del hígado y permiten la formación de urea a partir del amoníaco y el ácido aspártico. La urea es un subproducto menos t

Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (19)
Similar a Bioquimica (unidad iv)
Similar a Bioquimica (unidad iv) (20)
Más de José Isaac Rivero Castañeda
Más de José Isaac Rivero Castañeda (6)
Último
Último (20)
Bioquimica (unidad iv)
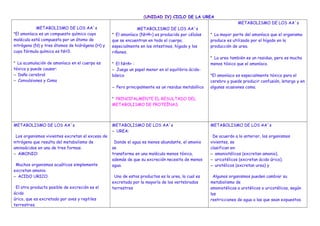
- 1. (UNIDAD IV) CICLO DE LA UREA METABOLISMO DE LOS AA´s *El amoníaco es un compuesto químico cuya molécula está compuesta por un átomo de nitrógeno (N) y tres átomos de hidrógeno (H) y cuya fórmula química es NH3. * La acumulación de amoníaco en el cuerpo es tóxica y puede causar: – Daño cerebral – Convulsiones y Coma METABOLISMO DE LOS AA´s * El amoníaco (NH4+) es producido por células que se encuentran en todo el cuerpo, especialmente en los intestinos, hígado y los riñones. * El NH4+ : – Juega un papel menor en el equilibrio ácido- básico – Pero principalmente es un residuo metabólico * PRINCIPALMENTE EL RESULTADO DEL METABOLISMO DE PROTEÍNAS METABOLISMO DE LOS AA´s * La mayor parte del amoníaco que el organismo produce es utilizado por el hígado en la producción de urea. * La urea también es un residuo, pero es mucho menos tóxico que el amoníaco. *El amoníaco es especialmente tóxico para el cerebro y puede producir confusión, letargo y en algunas ocasiones coma. METABOLISMO DE LOS AA´s Los organismos vivientes excretan el exceso de nitrógeno que resulta del metabolismo de aminoácidos en una de tres formas. – AMONIO: Muchos organismos acuáticos simplemente excretan amonio. – ACIDO URICO: El otro producto posible de excreción es el ácido úrico, que es excretado por aves y reptiles terrestres. METABOLISMO DE LOS AA´s – UREA: Donde el agua es menos abundante, el amonio se transforma en una molécula menos tóxica, además de que su excreción necesita de menos agua. Uno de estos productos es la urea, la cual es excretada por la mayoría de los vertebrados terrestres METABOLISMO DE LOS AA´s De acuerdo a lo anterior, los organismos vivientes, se clasifican en: – amoniotélicos (excretan amonio), – uricotélicos (excretan ácido úrico). – urotélicos (excretan urea) y Algunos organismos pueden cambiar su metabolismo de amoniotélicos a urotélicos o uricotélicos, según las restricciones de agua a las que sean expuestos.
- 2. CICLO DE LA UREA La urea: – Se sintetiza en el hígado – Por las enzimas del ciclo de la urea – Secretada al torrente sanguíneo – Filtrada en los riñones – Excretarse en la orina. CICLO DE LA UREA El ciclo de la urea – Fue encontrado en 1932 por Hans Krebs y Kurt Henseleit, – Fue el primer ciclo metabólico elucidado DEGRADACION DE LOS AA´s Los AA´s se interconvierten o degradan durante: – La eliminación de las proteínas celulares – La eliminación de las proteínas de la dieta – Ayuno prolongado DEGRADACION DE LOS AA´s * Etapas de la degradación – Separación del grupo amino: *TRANSAMINACIÓN *DESAMINACIÓN OXIDATIVA – Eliminación del grupo amino: *SÍNTESIS DE UREA TRANSAMINACION * Localización: – Citosol * Enzimas: – Transaminasas o aminotransferasas *Acción: – Catalizan la transferencia del grupo amino de un alfaaminoácido a un alfa-cetoácido * Sustratos y productos: – Alfa-aminoácido y alfa-cetoácido. TRANSAMINACION * Reacciones reversibles * También se usan para la síntesis de AA´s *Grupo prostético: – PLP (fosfato de piridoxal) * Alfa-cetoácido que recibe el grupo amino: – Alfa-cetoglutarato, que se transforma en GLUTAMATO – De esta manera todo el amoníaco es transferido al GLUTAMATO y de esta forma encaminado hacia la síntesis de urea.
- 3. – Degradación del esqueleto carbonado: *ALFA-CETO-ÁCIDOS intermediarios del Ciclo de Krebs DESAMINACION OXIDATIVA * Enzimas: – Glutamato deshidrogenasa y – Oxidasas * Acción: – Se remueve el grupo amino como amoniaco libre – Se acopla a una oxidación * Sustratos: – Alfa-aminoácido: GLUTAMATO * Productos: – Alfa-cetoácido: ALFA- CETOGLUTARATO DESAMINACION OXIDATIVA DESAMINACION OXIDATIVA * Reacción reversible * Grupo prostético: – NADH – NADPH * Localización: – Mitocondria – De esta manera el amoníaco es secuestrado en este compartimiento – Evitando que circule libremente. DEGRADACION DE LOS AA´s *El catabolismo del grupo amino de los AA ´s, se cumple en 3 procesos enzimáticos que ocurren en secuencia: – Transaminación – Desaminación oxidativa – Ciclo de la urea. CICLO DE LA UREA * Solo ocurre en hígado * Convierte amonio en urea *Menos tóxica * Parte ocurre en la mitocondria y parte en el citosol. CICLO DE LA UREA * Cinco reacciones – 3 citosol – 2 mitocondria * Transportadores para: – Citrulina – Ornitina * Paso limitante:
- 4. CICLO DE LA UREA *Carbamoil fosfato sintetasa I: – Localización: mitocondria – Función: síntesis de carbamoil fosfato – Sustrato: amonio – Requiere: 2 ATPs y bicarbonato – Producto: carbamoil fosfato. *Ornitina transcarbamilasa: – Localización: mitocondria – Función: condensa el carbamoil fosfato con la ornitina – Sustrato: carbamoil fosfato – Producto: citrulina CICLO DE LA UREA * Arginosuccinato sintasa: – Localización: citoplasma – Función: condensa el aspártico con la citrulina – Requiere: ATP – Sustrato: citrulina – Producto: ácido arginosuccínico – El segundo nitrógeno necesario para la formación de la urea lo aporta el ácido aspártico. – Síntesis de CARBAMOIL FOSFATO (CPS I). * Enzimas – Carbamoil fosfato sintetasa I – Ornitina transcarbamilasa – Arginosuccinato sintasa – Arginosuccinato liasa – Arginasa. ORIGEN DEL ASPARTATO PARA LA REACCION DE LA ARGINOSUCCINATO SINTETASA. EL CICLO DE LA UREA ESTA LIGADO AL ACIDO CITRICO CICLO DE LA UREA CICLO DE LA UREA CICLO DE LA UREA
- 5. • Arginosuccinato liasa: – Localización: citoplasma – Función: el ácido arginosuccínico formado en la reacción anterior se divide en fumárico y arginina. – Sustrato: ácido arginosuccínico – Producto: arginina. • Arginasa: – Localización: citoplasma – Función: en esta reacción se forma urea después de la hidrólisis de la arginina – Sustrato: arginina – Producto: UREA y ornitina (que pasará del citoplasma a la mitocondria, donde comienza de nuevo el ciclo) TRANSPORTE DE AMONIACO HASTA EL HIGADO * El amonio cuando está en tejido se une al ácido glutámico y forma la glutamina, * Reacción mediada por la glutamina sintasa * La glutamina pasa a la circulación, donde es *Capturada por el hígado * Convierte de nuevo en ácido glutámico *Reacción mediada por la glutaminasa. 2 NH4 + HCO3 + 3 ATP UREA + 2ADP + 4 Pi + AMP + 5 H. TRANSPORTE DE AMONIACO HASTA EL HIGADO En nuestro organismo, la glutamina, es el aminoácido encargado de: – Almacenar de manera temporal, – Mantener dentro de los niveles aceptados y – Transportar el amonio. DESTINO DEL ESQUELETOS CARBONADOS DE LOS AMINOACIDOS El catabolismo de los aminoácidos, requiere una vía exclusiva para el metabolismo del esqueleto del carbono de los aminoácidos. Sus productos finales son intermediarios del Ciclo de Krebs. *AA´s formadores de: – Piruvato – Acetil-CoA – Aceto-acetil-CoA – Alfa-cetoglutarato
- 6. – Succinil-CoA – Fumarato – Oxaloacetato AA´s FORMADORES DE PIRUVATO * Puede usarse para formar OXALOACETATO (gluconeogénesis) o ACETIL-CoA e ingresar al Ciclo de Krebs. AA´s FORMADORES DE ACETIL-CoA *leuicina *isoleucina * triptófano. AA´s FORMADORES DE ACETO-ACETIL-CoA *leucina * lisina *fenilalanina *tirosina *triptófano. AA´s FORMADORES DE ALFACETOGLUTARATO *arginina *histidina *prolina *glutamina *acido AA´s FORMADORES DE SUCCINIL-CoA *isoleucina *metionina *valina. AA´s FORMADORES FUMARATO *fenilalanina *tirosina. AA´s FORMADORES DE OXALOACETATO
- 7. DEGRADACION DE LOS AA´s *GLUCONEOGÉNICOS – Permiten la síntesis de glucosa – Los que dan origen a: ---- Piruvato ----alfa-cetoglutarato ----Succinil-CoA ------ Fumarato ------Oxaloacetato *CETOGÉNICOS – Permiten la síntesis de cuerpos cetónicos – Los 6 AA's que originan Acetil-CoA y Acetoacetol-CoA < Ile < Leu < Trp < Phe < Tyr glutamico NOTA: El alfa-cetoglutarato es esencial en el proceso de transaminación. *Asparagina * Acido aspartico.
- 8. < Lys DEGRADACION DE LOS AA´s *AA´s GLUCONEOGÉNICOS Y CETOGÉNICOS – Ile – Trp – Phe – Tyr COMPLEJO DE LA ACIDO GRASO SINTETASA *Ocurre en el CITOSOL a partir de ACETIL-CoA *Ocurre en situacion de alimentacion. *Enzima multifuncional que consiste en dos cadenas Idénticas * Se llevan a cabo 7 reacciones *Todas las enzimas de la via, excepto la primera, se agrupan en un complejo multienzimatico denominado Acido Graso Sintetasa (AGS) COMPLEJO DE LA ACIDO GRASO SINTETASA * Se encuentra asociado al Reticulo Endoplasmico Liso *En los mamiferos esta constituida por una sola cadena de aminoacidos, que tiene sitios activos para todas las actividades enzimaticas del complejo. * Esta cadena incluye un dominio acarreador de acilos, llamado ACP (acyl-carrier protein) SINTETASA DE ACIDOS GRASOS Enzimas: – Acetil-CoA Transacilasa – Malonil-CoA Transacilasa – 3-Cetoacil-ACP Sintasa (Enzima condensante) – 3-Cetoacil-ACP Reductasa – D-3-Hidroxiacil-ACP Deshidratasa – Enoil-ACP Reductasa – ACP
- 9. COMPLEJO DE LA ACIDO GRASO SINTETASA * Grupos prostéticos de la AGS incluyen: – El grupo tiol (-SH) de la cadena lateral de un residuo de cisteina en el dominio de la enzima condensante (es el que une el acetil-CoA inicial). – El grupo tiol (-SH) de la fosfopanteteina, que se une covalentemente a la ACP (es el que une el Malonil-CoA) CARBOXILACION DEL ACETIL-CoA llll ACIDO GRASO SINTASA
- 10. ENZIMA: ACETIL-CoA CARBOXILASA COMPLEJO DE LA ACIDO GRASO SINTETASA * Grupos prostéticos de la AGS incluyen: – El grupo tiol (-SH) de la cadena lateral de un residuo de cisteina en el dominio de la enzima condensante (es el que une el acetil-CoA inicial). – El grupo tiol (-SH) de la fosfopanteteina, que se une covalentemente a la ACP (es el que une el Malonil-CoA) SINTETASA DE ACIDOS GRASOS Enzimas: – Acetil-CoA Transacilasa – Malonil-CoA Transacilasa – 3-Cetoacil-ACP Sintasa (Enzima condensante) – 3-Cetoacil-ACP Reductasa – D-3-Hidroxiacil-ACP Deshidratasa – Enoil-ACP Reductasa ACETIL-CoA TRANSACILASA Transfiere el Acetilo de la Coenzima A, al grupo -SH del residuo de cisteina (grupo prostetico).
- 11. – ACP MALONIL-CoA TRANSACILASA Transfiere el malonil-CoA a la ACP (Proteina Acarreadora de Acilos) que tiene como grupo prostetico el acido Pantotenico. BETA-CETOACIL-ACP SINTASA o ENZIMA CONDENSANTE Forma Acetoacetil-ACP: Transfiriendo el Acetilo al Carbono 2 de Malonil- ACP, Con la eliminacion simultanea del radical carboxilo del Malonil, en forma de CO2. BETA-CETOACIL-ACP REDUCTASA D-3-HIDROXIACIL-ACP DESHIDRATASA
- 12. Esta enzima reduce en forma estereoespecifica, el 3-Cetoacido a un 3-Hidroxiacido, formado unicamente el isomero D, isomero contrario al de la beta-oxidacion. El NADPH que se necesita para la reduccion puede provenir de la via de las Pentosas o de la actividad de la Enzima Malica. La enzima es estereoespecifica A partir del D-Hidroxiacido forma exclusivamente el trans-Δ2-Enoil-ACP
- 13. ENOIL-ACP REDUCTASA Reduce el enlace doble del Enoil-ACP para formar el Acil-ACP saturado. El Acil-ACP producido toma el lugar de la Acetil-ACP para repetir el ciclo de reacciones, de esta manera en cada ciclo de sintesis, se anaden dos carbones a la cadena, donados por Malonil-CoA. SINTETASA DE ACIDOS GRASOS SINTETASA DE ACIDOS GRASOS La secuencia de reacciones de la AGS se repite unicamente 7 veces, por cada molecula sintetizada. * Por lo tanto, el complejo solo puede sintetizar Palmitil- ACP. * La sintesis de una molecula de Palmitato consume: – 8 moleculas de Acetil-CoA, – 7 de ATP y – 14 de NADPH.
- 14. Para sintetizar el acido graso C16 se necesitan: – 8 moleculas de Acetil-CoA (16/2) – Para participar en la sintesis siete de estas moleculas deben convertirse en Malonil-CoA – Lo cual consume 7 moleculas de ATP. * Ademas, en cada paso por el ciclo de sintesis se consumen: – 2 moleculas de NADPH, para reducir los intermediarios.
- 15. METABOLISMO DE LA GLUCOSA * La glucosa debe entrar a la célula y quedar atrapada allí para poder ser metabolizada * Su ingreso a la célula depende de la presencia de transportadores específicos * La insulina y el glucagon regulan el ingreso de glucosa a las células * La glucosa se debe oxidar para obtener energía. – Todos los organismos hacen glicólisis – Humanos: todos los tejidos hacen glicólisis *La célula también esta en capacidad de sintetizar glucosa. – Objetivo: mantener un suministro constante de glucosa para los órganos cuyo combustible es este. Pag. 5 GLUCONEOGENESIS * Síntesis de glucosa a partir de sustratos NO CARBOHIDRATOS * Se realiza principalmente en hígado. * También se realiza aunque en menor grado en: – Corteza renal – Intestino delgado * La vía completa se realiza en: – mitocondria y – citosol GLUCONEOGENESIS *Su objetivo es mantener la glicemia – Existen órganos cuyo combustible es la glucosa: < Cerebro < Glóbulos rojos < Médula renal < Cristalino y córnea – Esto obliga a mantener los valores de glicemia dentro de límites normales, a pesar del ayuno < Requiere energía. GLUCONEOGENESIS < La gluconeogénesis transcurre a través de una serie de reacciones comunes con la glicólisis, en sentido inverso < En cierto modo se puede considerar como el reverso de la glicólisis < Existen 3 reacciones de la glicólisis muy alejadas del equilibrio: – Son irreversibles – Existen reacciones de desvío para sortear estas Reacciones.
- 16. REACCIONES IRREVERSIBLES DE LA GLICOLISIS GLUCONEOGENESIS Sustratos – Piruvato – Glicerol – Lactato (músculo y glóbulos rojos) – AA's provenientes del catabolismo proteico (músculo) – Acidos Grasos NO SON SUSTRATOS PARA LA GLUCONEOGENESIS * Sustratos a nivel renal – Lactato – Piruvato – Glicerol – Glutamina * Sustratos a nivel hepático – Lactato – Piruvato – Glicerol – Alanina nota: <7 de las enzimas usadas en la vía glicolítica, son usadas en la gluconeogénesis < Las 3 reacciones restantes son irreversibles <En estos puntos se requieren reacciones de desvío
- 17. PRIMERA PARTE: Formación de Oxaloacetato a partir de Piruvato PRIMERA REACCIÓN DE DESVIO: FORMACIÓN DE PEP A PARTIR DE PIRUVATO < No se puede realizar en un solo paso: – Primera parte: mitocondria – Segunda parte: citosol – Se requieren sistemas de transporte: MMI < Requiere dos reacciones: – Formación de OA a partir de piruvato – Formación de PEP a partir de OA. SEGUNDA PARTE: Formación de PEP a partir de OA <Enzima: – PEP carboxiquinasa – Requiere GTP – Descarboxilación del oxalacetato hasta PEP – Citosólica. Pg17 PRIMERA PARTE: Formación de Oxaloacetato a partir de Piruvato <Enzima: – Piruvato carboxilasa – Requiere ATP – Cataliza la Carboxilación del piruvato hasta Oxalacetato – Localización: Mitocondrial – El OA sale como Malato
- 18. Entre el: – PEP y la fructosa -1,6-BP todas las reacciones son comunes a la glicólisis pues estas son libremente reversibles. TERCERA REACCION DE DESVIO: Formación de F-6-P <Enzima – Fructosa-1,6- bifosfatasa – Cataliza la fosforólisis del fosfato en la posición 1 de la fructosa. SUSTRATOS GLUCONEOGENICOS Formación de GLUCOSA LIBRE Enzima: – Glucosa -6-fosfatasa – Está presente en hígado y riñón – Permite liberar la glucosa a la sangre.
- 19. SUSTRATOS GLUCONEOGENICOS PRIMERA PARTE DE LA GLICÓLISIS
- 20. Pag.28 PRIMERA PARTE DE LA GLICÓLISIS < Se forma una hexosa fosforilada < El producto son dos triosas fosforiladas, isómeros entre si < Las triosas se interconvierten entre si y 2 Gliceraldehido-3-P pasan a la siguiente fase de la glicólisis <Se consumen dos ATP SEGUNDA PARTE DE LA GLICÓLISIS Se oxidan 2 Gliceraldehido-3-P hasta ácido pirúvico. < Se producen – 2 NADH – 4 ATP < El destino del piruvato depende de las condiciones redox de lacélula. CICLOS de CORI y de la GLUCOSA- ALANINA <Existen 2 ciclos entre tejidos, en los que esta involucrada la gluconeogénesis. < Estos ciclos consisten en: – Gluconeogénesis hepática, seguida de – Glicólisis en un tejido periférico < Objetivo: – Proporcionar un mecanismo para suministrar glucosa continuamente a los tejidos que dependen de la glucosa como fuente primaria de energía. CICLOS de CORI y de la GLUCOSA-ALANINA <Cuales son las condiciones para que estos ciclos funcionen: – Solo son funcionales en aquellos tejidos en los que no se oxida la glucosa completamente hasta CO2 y H2O – El tejido periférico debe eliminar como productos de la glicólisis: < Lactato o < Alanina
- 21. Diferencias entre los CICLOS de cori y glucosa- ala Cori < El intermediario de 3 carbonos que es reciclado: LACTATO < Tejidos involucrados: aquellos que pueden producir lactato como producto final – Eritrocitos – Músculo y – En general cualquier tejido < El NADH producido por la glicólisis, es utilizado para reducir el piruvato hasta lactato. < Consumo neto de ATP para realizar gluconeogénesis: 4 Glucosa-alanina < El intermediario de 3 carbonos que es reciclado: ALANINA < Tejidos involucrados: aquellos que pueden producir alanina como producto final – Músculo – Eritrocitos, no liberan alanina < El NADH producido por la glicólisis, no puede ser utilizado para reducir el piruvato hasta lactato. < Esto implica: – El uso de las lanzaderas en los tejidos con mitocondrias y – La presencia de O2 < El uso de alanina implica el funcionamiento simultáneo del ciclo de la urea < Consumo neto de ATP para realizar gluconeogénesis: 2-4 CICLO de CORI <El ciclo de Cori involucra la utilizacion de lactato, producido en la glucolisis en tejidos no-hepaticos, (como el musculo y los eritrocitos) como una fuente de carbono para la gluconeogenesis hepática. < En esta forma el hígado puede convertir el producto de la glucólisis anaerobia, lactato, otra vez en glucosa para su reutilización por parte de tejidos no-hepaticos.
- 22. CICLO de la ALANINA-GLUCOSA El ciclo glucosa-alanina es utilizado como mecanismo para que el músculo esqueletico elimine nitrógeno. < La oxidación de la glucosa produce piruvato que puede ser transaminado a alanina < Esta reacción es catalizada por la alanino transaminasa, ALT (transaminasa glutamato- piruvato o SGPT) < La alanina ingresa al torrente sanguíneo y es transportado al hígado Regulación de la Gluconeogénesis La regulacion de la gluconeogenesis estará en contraste directo a la regulación de la glicólisis. < En general, los efectores negativos de la glicólisis son efectores positivos de la gluconeogénesis. < El sitio mas significativo para controlar el flujo hacia la oxidación o la síntesis de glucosa, es la regulación de la actividad de: – la PFK-1 en la glicólisis y – la F-1,6-BPasa en la gluconeogénesis – El efector alostérico más importante: Regulación de la Gluconeogénesis CICLO de la ALANINA-GLUCOSA En el hígado, la alanina es convertida en piruvato que es utilizado como fuente de carbono para la gluconeogenesis. < La glucosa nueva que ha sido formada puede entonces entrar a la sangre para regresar al musculo < Durante períodos de ayuno, la proteína del musculo esqueletico se degrada por el valor energético de los carbonos de los aminoácidos y la alanina es el principal aminoacido de esa proteína. < El grupo amino transportado desde el músculo al hígado en la forma de alanina se convierte en urea en el ciclo de la urea y es excretado. Control local: – Incluye regulación alostérica recíproca por nucleotidos de adenina. < PFK-1 (glicólisis): – Inhibida por ATP – Estimulada por AMP < F-1,6-BPasa (Gluconeogénesis) – Inhibida por AMP – Esto asegura que: < Cuando el ATP esta alto (AMP debería estar bajo):
- 23. < FRUCTOSA-2,6-BIFOSFATO – la glucosa no se degrada para obtener ATP – la glucosa se guarda como glucógeno < Cuando el ATP esta bajo (AMP debería estar alto): – la célula no gasta energía en sintetizar glucosa. Control global: – Incluye los efectos recíprocos de la cascada del cAMP, que son desencadenadas por el glucagon: < La fosforilación de enzimas y proteínas regulatorias por la PKA (Protein Kinasa dependiente de cAMP) resulta en la inhibición de la glicólisis y la estimulación de la gluconeogenesis – Las proteínas relevantes para esta vía son: <Piruvato quinasa < CREB (cAMP Response Element Binding protein) < La enzima bifuncional (PFK-2/F-2,6-BPasa) <Piruvato quinasa: – es inhibida cuando se fosforila <CREB (cAMP Response Element Binding protein): – al ser fosforilado, activa la transcripción del gen de la PEP carboxiquinasa. < La enzima bifuncional (PFK-2/F-2,6-BPasa): – permite la síntesis y degradación del regulador alostérico fructosa-2,6-bisfosfato. – F-2,6-BPasa: activa cuando esta fosforilada. – PFK-2: activa cuando esta desfosforilada. < La PFK-2/F-2,6-BPasa es fosforilada por la protein quinasa A (PKA), que es una quinasa dependiente del cAMP <Cuando la F-2,6-BPasa esta activa (forma fosforilada de la enzima bifuncional), el resultado es: < Actividad: Fosfofructoquinasa-2 (PFK-2) y < Cataliza la síntesis de la F-2,6-BP por la fosforilación de la fructosa-6-fosfato – Forma FOSFORILADA de la enzima: < Actividad: Fructosa-2,6-bifosfatasa (F-2,6- BPasa) e < Hidroliza la F-2,6-BP a fructosa-6-fosfato y P Regulación de la Gluconeogénesis
- 24. – La actividad de la F-1,6-BFasa es fuertemente estimulada y – La actividad de la PFK-1 esta fuertemente inhibida. – Por lo tanto esta activa vía gluconeogénica < El resultado metabólico de la fosforilación de la enzima bifuncional es: – La estimulación alosterica de la PFK-1 se detiene – La inhibición alosterica de la F-1,6-BPasa se elimina, y – El flujo neto es gluconeogénico METABOLISMO DE LA GLUCOSA 1)Degradación de la glucosa – Glucólisis < Sitio: – Anaerobia: citosol – Aerobia: Citosol + Mit < Sustrato: – Glucosa < Producto: – Anaerobia: Piruvato/Lactato – Aerobia: CO2 + H2O 2)Síntesis de glucosa – Gluconeogénesis < Sitio: – Citosol + Mit < Sustrato: – Piruvato – Lactato – Glicerol – AA's < Producto – Glucosa REGULACION DE LA GLICEMIA < Insulina < Glucagon – Células b-pancreas -Células a-pancreas – Producción: -Producción: < Incremento glicemia <Disminución glicemia -Efecto: –Efecto: 1)salida de 1)Entrada de glucosa a glucosa a la sangre la célula. )2 Disminución de la 2)Aumento de la glicemia Glicemia