DANIELA BAEZA ZAVALA
GABRIELCABRERA HERRERA
DIANA LAURA JACOME GARCIA
JOSÉ NAJIB RICÁRDEZ RODRÍGUEZ
Sistema del
Universidad Veracruzana
Inmunología
2.
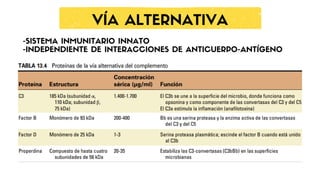
QUE ES ELSISTEMA DE
COMPLEMENTO:
Conjunto de mas de
50 proteínas
plasmáticas que
actúan eliminando
patógenos, células
dañadas o
inmunocomplejo.
Las proteínas del
complemento son
proteínas plasmáticas
normalmente
inactivas
Se activan solo en
condiciones
particulares para
generar productos
que median varias
funciones efectoras
3.
CARACTERISTICAS:
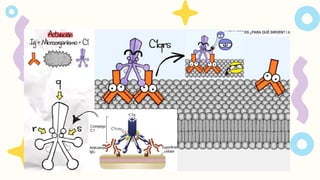
Al sistema del
complementolo activan
microbios y anticuerpos y
lectinas unidos a los
microbios y otros
antígenos. La activación del
complemento implica la
proteolisis secuencial de
proteínas para generar
complejos enzimáticos con
actividad proteolítica
ZIMÓGENOS
4.
CARACTERISTICAS:
Varios de losproductos
escindidos del complemento con
actividad biológica se unen de
forma covalente a las
superficies microbianas, a
anticuerpos unidos a los microbios
y a otros antígenos, y a cuerpos
apoptósicos.
La activación del
complemento la inhiben
proteínas reguladoras
presentes en las células
normales del hospedador y
que faltan en los microbios
5.
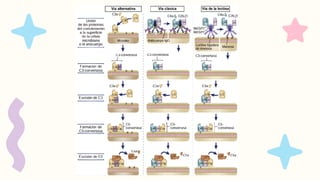
CARACTERISTICAS:
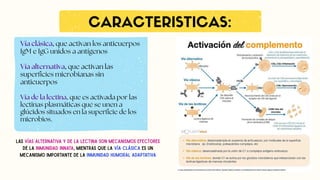
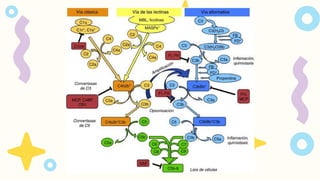
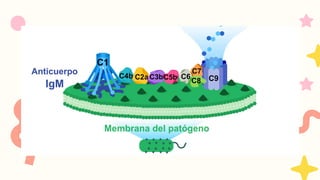
Vía clásica, queactivan los anticuerpos
IgM e IgG unidos a antígenos
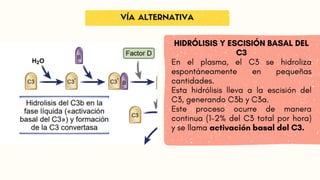
Vía alternativa, que activan las
superficies microbianas sin
anticuerpos
Vía de la lectina, que es activada por las
lectinas plasmáticas que se unen a
glúcidos situados en la superficie de los
microbios.
Las vías alternativa y de la lectina son mecanismos efectores
de la inmunidad innata, mientras que la vía clásica es un
mecanismo importante de la inmunidad humoral adaptativa
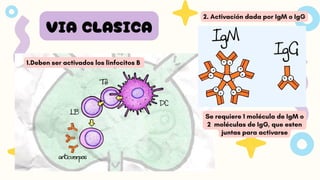
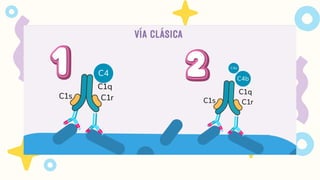
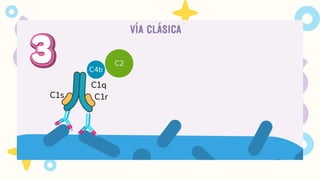
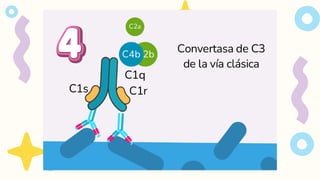
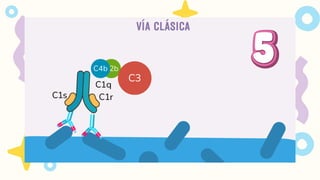
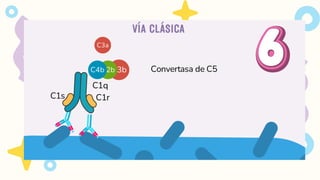
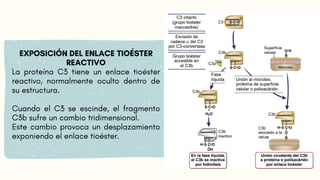
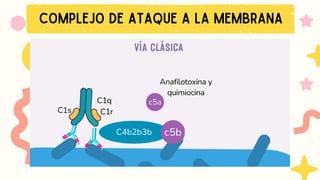
VIA CLASICA
1.Deben seractivados los linfocitos B
2. Activación dada por IgM o IgG
Se requiere 1 molécula de IgM o
2 moléculas de IgG, que esten
juntas para activarse
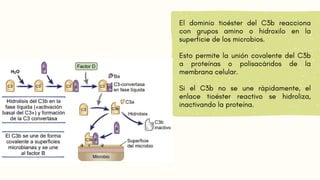
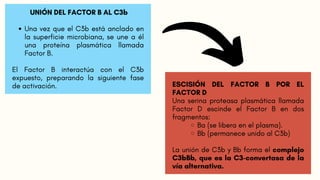
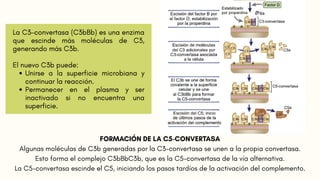
FORMACIÓN DE LAC5-CONVERTASA
Algunas moléculas de C3b generadas por la C3-convertasa se unen a la propia convertasa.
Esto forma el complejo C3bBbC3b, que es la C5-convertasa de la vía alternativa.
La C5-convertasa escinde el C5, iniciando los pasos tardíos de la activación del complemento.
24.
La Properdina esuna proteína reguladora del complemento que
estabiliza la C3-convertasa (C3bBb) y prolonga su actividad.
Es la única proteína reguladora positiva de la vía alternativa, lo que
significa que favorece y potencia la activación del complemento en lugar
de inhibirlo.
PAPEL DE LA PROPERDINA
1.Se une al complejo C3bBb (C3-convertasa de la vía alternativa).
2. Evita su degradación rápida, prolongando su actividad.
3.Favorece la generación de más C3b, amplificando la respuesta inmune.
Sin la Properdina, la C3-convertasa es inestable y se degrada en poco tiempo.
Se une preferentemente a las superficies microbianas, pero no a células del hospedador-.proteínas
reguladoras (ej. Factor H y Factor I)
25.
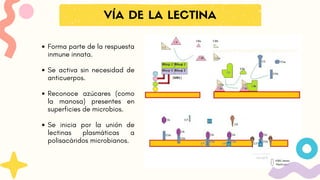
VÍA DE LALECTINA
Forma parte de la respuesta
inmune innata.
Se activa sin necesidad de
anticuerpos.
Reconoce azúcares (como
la manosa) presentes en
superficies de microbios.
Se inicia por la unión de
lectinas plasmáticas a
polisacáridos microbianos.
26.
VÍA DE LALECTINA
Proteínas lectinas involucradas:
Lectina ligadora de manosa (Mannose-Binding Lectin) y ficolinas son proteínas solubles en
plasma.
Estructuralmente son similares a C1q del complemento clásico.
Tipos de lectinas:
MBL, ficolina L y ficolina H → plasmáticas.
Ficolina M → secretada principalmente por macrófagos activados.
Estructura:
Ambas poseen:
Dominio amino terminal similar al colágeno → permite formar estructuras helicoidales
triples.
Dominio carboxilo terminal:
MBL → tipo lectina, específica para manosa.
Ficolinas → tipo fibrinógeno, reconoce N-acetilglucosamina.
Estas estructuras permiten la formación de oligómeros y la interacción con patógenos.
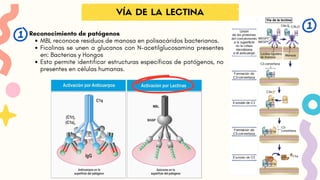
Reconocimiento de patógenos
MBLreconoce residuos de manosa en polisacáridos bacterianos.
Ficolinas se unen a glucanos con N-acetilglucosamina presentes
en: Bacterias y Hongos
Esto permite identificar estructuras específicas de patógenos, no
presentes en células humanas.
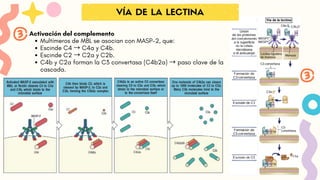
VÍA DE LA LECTINA
29.
VÍA DE LALECTINA
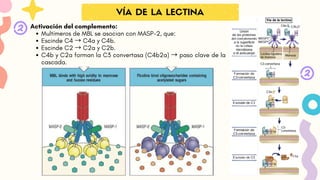
Activación del complemento:
Multímeros de MBL se asocian con MASP-2, que:
Escinde C4 → C4a y C4b.
Escinde C2 → C2a y C2b.
C4b y C2a forman la C3 convertasa (C4b2a) → paso clave de la
cascada.
30.
Activación del complemento
Multímerosde MBL se asocian con MASP-2, que:
Escinde C4 → C4a y C4b.
Escinde C2 → C2a y C2b.
C4b y C2a forman la C3 convertasa (C4b2a) → paso clave de la
cascada.
VÍA DE LA LECTINA
31.
Etapas siguientes
Desde laformación de la C3 convertasa, la vía continúa igual que
la vía clásica:
Escisión de C3 → C3a (inflamación) y C3b (opsonización).
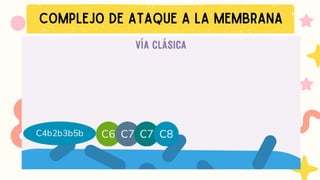
Formación de C5 convertasa.
Activación del complejo de ataque a membrana (MAC): C5b-C9
→ lisis del patógeno.
VÍA DE LA LECTINA
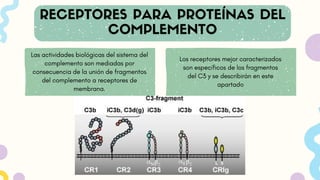
Las actividades biológicasdel sistema del
complemento son mediadas por
consecuencia de la unión de fragmentos
del complemento a receptores de
membrana.
Los receptores mejor caracterizados
son específicos de los fragmentos
del C3 y se describirán en este
apartado
39.
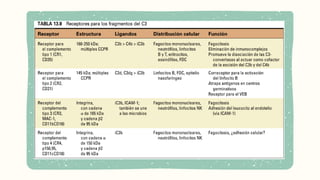
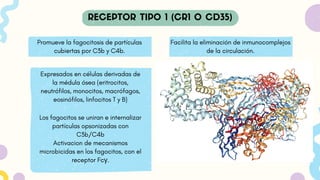
Promueve la fagocitosisde partículas
cubiertas por C3b y C4b.
Expresados en células derivadas de
la médula ósea (eritrocitos,
neutrófilos, monocitos, macrófagos,
eosinófilos, linfocitos T y B)
Facilita la eliminación de inmunocomplejos
de la circulación.
Los fagocitos se uniran e internalizar
partículas opsonizadas con
C3b/C4b
Activacion de mecanismos
microbicidas en los fagocitos, con el
receptor Fcγ.
40.
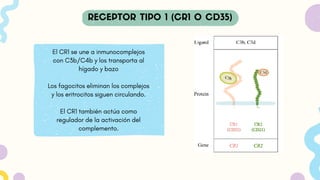
El CR1 seune a inmunocomplejos
con C3b/C4b y los transporta al
hígado y bazo
Los fagocitos eliminan los complejos
y los eritrocitos siguen circulando.
El CR1 también actúa como
regulador de la activación del
complemento.
41.
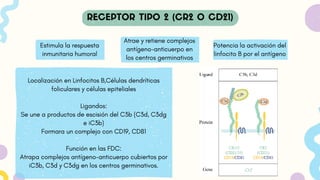
Estimula la respuesta
inmunitariahumoral
Localización en Linfocitos B,Células dendríticas
foliculares y células epiteliales
Ligandos:
Se une a productos de escisión del C3b (C3d, C3dg
e iC3b)
Formara un complejo con CD19, CD81
Función en las FDC:
Atrapa complejos antígeno-anticuerpo cubiertos por
iC3b, C3d y C3dg en los centros germinativos.
Atrae y retiene complejos
antígeno-anticuerpo en
los centros germinativos
Potencia la activación del
linfocito B por el antígeno
42.
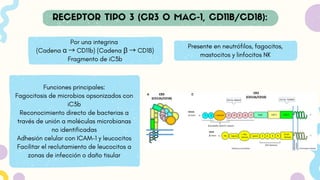
Por una integrina
(Cadenaα → CD11b) (Cadena β → CD18)
Fragmento de iC3b
Funciones principales:
Fagocitosis de microbios opsonizados con
iC3b
Reconocimiento directo de bacterias a
través de unión a moléculas microbianas
no identificadas
Adhesión celular con ICAM-1 y leucocitos
Facilitar el reclutamiento de leucocitos a
zonas de infección o daño tisular
Presente en neutrófilos, fagocitos,
mastocitos y linfocitos NK
43.
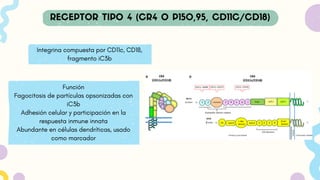
Integrina compuesta porCD11c, CD18,
fragmento iC3b
Función
Fagocitosis de partículas opsonizadas con
iC3b
Adhesión celular y participación en la
respuesta inmune innata
Abundante en células dendríticas, usado
como marcador
44.
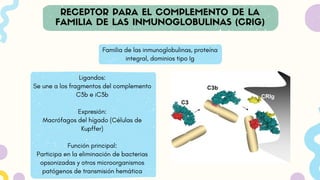
Familia de lasinmunoglobulinas, proteína
integral, dominios tipo Ig
Ligandos:
Se une a los fragmentos del complemento
C3b e iC3b
Expresión:
Macrófagos del hígado (Células de
Kupffer)
Función principal:
Participa en la eliminación de bacterias
opsonizadas y otros microorganismos
patógenos de transmisión hemática
45.
La activación delsistema de complemento es
altamente regulada para evitar daño a las
células propias del organismo
REGULACIÓN DE LA ACTIVACIÓN
DEL COMPLEMENTO
Esto es especialmente importante porque los
productos del complemento tienen una alta
capacidad destructiva.
46.
Evita la destrucciónde células propias
Controla la inflamación y la lisis celular
Regulación se da en distintas fases
En la activación de C3 y C5 convertasa: los
puntos críticos donde se forma el complejo
que activa las siguientes etapas del sistema
de complemento.
En la fase terminal: donde se forma el
complejo de ataque a membrana (MAC).
47.
PROTEÍNAS REGULADORAS DELCOMPLEMENTO (EJEMPLOS):
C1-INH (Inhibidor de C1): regula la
vía clásica al inhibir la actividad de
C1r y C1s
CD59 (Protectina): bloquea la
formación del MAC en células
propias.
Factor I: proteasa que inactiva C3b
y C4b
Factor H: cofactor del Factor I para
la degradación de C3b; compite
con el Factor B para unirse a C3b
CD55 (DAF – Decay Accelerating
Factor): descompone las
convertasas C3 y C5.
Vitronectina y clústerina: impiden la
inserción del MAC en la membrana
celular.
48.
FUNCIONES DEL COMPLEMENTO
Lasprincipales funciones del sistema del complemento en la inmunidad
innata y en la inmunidad humoral adaptativa son promover la fagocitosis
de los microbios sobre los cuales se activa el complemento, estimular la
inflamación e inducir la lisis de estos microbios.
49.
FUNCIONES DEL COMPLEMENTO
Estimulaciónde la Respuesta Inflamatoria:
Algunos fragmentos del complemento
actúan como anafilotoxinas:
C3a, C4a y C5a
Estos fragmentos activan:
Mastocitos → liberan histamina
(vasodilatación).
Neutrófilos → migración y activación.
Células endoteliales → aumento de
permeabilidad vascular.
Resultado: Inflamación aguda,
reclutamiento de más células inmunitarias.
Opsonización y Fagocitosis:
El complemento marca microbios con
proteínas como C3b, iC3b y C4b.
Estas proteínas se fijan a la superficie
de los patógenos tras la activación del
complemento.
Macrófagos y neutrófilos reconocen
estas marcas mediante receptores
específicos.
Resultado: Fagocitosis rápida y eficiente
del microorganismo.
*Opsonización = “hacer comestible” al
patógeno para los fagocitos.
50.
FUNCIONES DEL COMPLEMENTO
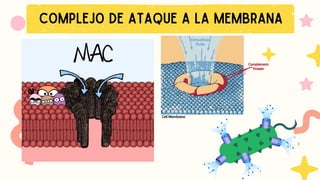
CitólisisMediada por el Complemento:
El MAC (Complejo de Ataque a la
Membrana) se forma al final de la
cascada del complemento.
MAC perfora la membrana del
microorganismo → lisis celular.
Es eficaz contra microorganismos con:
Membranas delgadas, como las
Neisseria spp.
Algunos microbios (con cápsulas gruesas)
resisten el ataque del MAC.
Eliminación de Complejos Inmunes:
Las proteínas del complemento se unen
a los complejos antígeno-anticuerpo.
Esto facilita su:
Solubilización.
Reconocimiento y eliminación por
fagocitos.
Es una forma de evitar que estos
complejos se depositen en tejidos y
causen daño.
51.
Vía Clásica
Afecta C1q,C1r, C4, C2 y C3.
C2 es la deficiencia más común.
Relacionada con lupus y
eliminación deficiente de
inmunocomplejos.
C3 es clave para opsonización; su
deficiencia causa infecciones
graves.
DEFICIENCIAS DEL COMPLEMENTO
Vía Alternativa
Factor B y D: infecciones
meningocócicas.
Properdina (ligada al X): mayor
riesgo de infecciones severas.
Fase Final
C5-C9 : infecciones por Neisseria
meningitidis y gonorrhoeae.
52.
Evasión del sistemadel complemento reclutando proteínas
reguladoras del complemento del hospedador
EVASIÓN DEL COMPLEMENTO POR MICROBIOS
Producción de proteínas que inhiben diferentes pasos
de la activación del complemento
Mediante la inhibición de la inflamación, mediado
por productos microbianos