Descargado 97 veces













![Componente C3 serico,
contiene un enlace tioester
1
Hidrolisis para
producir
2
3
Las membranas de los mamíferos contienen [ ]
altas de ácido siálico = desactivación de C3b
4
Factor B
5
Factor
B
Factor
D
6
Factor
B
Factor
D
BaBb](https://image.slidesharecdn.com/sistemadelcomplemento-161208043402/85/Sistema-del-complemento-libro-KUBY-18-320.jpg)






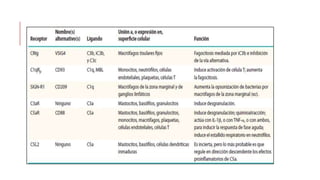












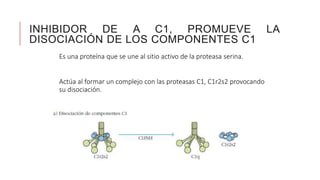


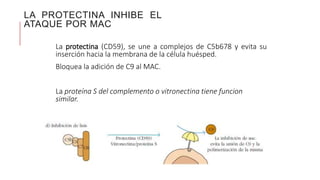







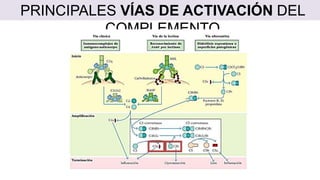
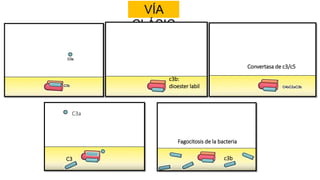
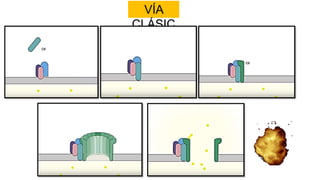
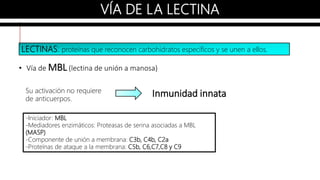
El sistema del complemento es un grupo de proteínas séricas que cooperan con el sistema inmunitario innato y adaptativo para eliminar agentes patógenos de la sangre y los tejidos. Interactúan entre sí en cascadas catalíticas e incluyen componentes como C1q, C3 y C5 que opsonizan bacterias para fagocitosis, inducen respuestas inflamatorias, y forman el complejo de ataque a la membrana. El sistema del complemento está regulado por proteínas como el factor I y la protectina para prevenir daño a cél












































































