Descargado 97 veces







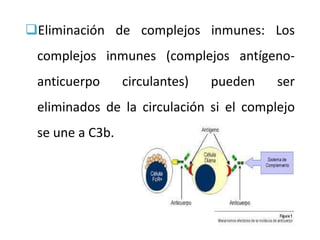


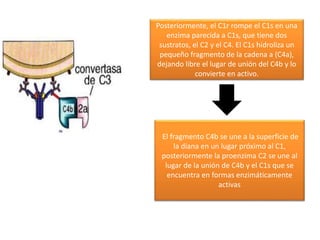
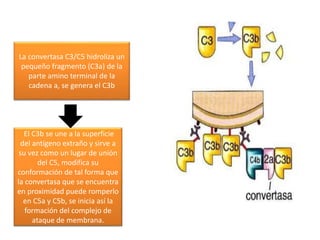












El sistema del complemento consta de proteínas plasmáticas que potencian la respuesta inflamatoria, facilitan la fagocitosis y dirigen la lisis celular. Se activa por tres vías: la clásica, la alterna y la de las lectinas, las cuales convergen en la formación del complejo de ataque a la membrana para destruir la célula diana.



































































