Descargado 124 veces








































































































![106
Los anteridios y arquegonios generalmente forman pequeños discos o platillos largamente
pedunculados Los esporangios carecen de casquetes y dan esporas asexuales. La reproducción es
alternada, como la de los musgos, pero también pueden reproducirse por pequeñas granulaciones o
progágulas que se forman en la cara superior de las hojas y que al ser arrastradas por e] agua dan
origen a una nueva planta El nombre proviene de la palabra griega hépar, hépatos = hígado, y se las
llama así porque en otros tiempos se empleaban para combatir las enfermedades del hígado.
Especies principales.
La Marchantia o Hepática de las
fuentes, sin tallo son muy comunes en
las grutas y en las rocas húmedas. Son
dioicas.
Las Jungermanias, con tallo hueco y
provisto de una doble serie de hojas sin
nervaduras; la cápsula se abre por 4
valvas.
Utilidad. — Contribuyen a aumentar la
capa de tierra vegetal en los bosques y
ayudan a llenar las turberas.
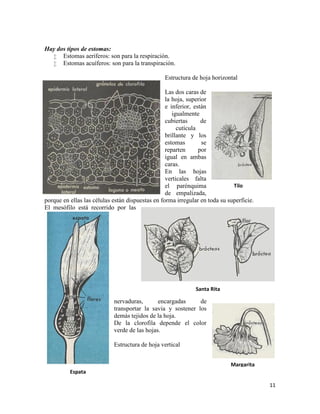
DIVISIÓN PTERIPOFITAS
Pteridofitas o criptógamas vasculares
Las pteridofitas se caracterizan por tener vasos destinados a la conducción de la savia; además
poseen raíces, tallos y hojas, y como tienen clorofila, son plantas autótrofas. La reproducción es
alternante.
Lo comprenden 3 clases:
Filicíneas o helechos con ramificación lateral
Equisetáceas, con ramificación verticilada.
Licopodáceas, con ramificación dicotómica
Por ser los helechos la clase más numerosa, han dado el nombre al subtipo (pteris = helecho).
HELÉCHOS VERDADEROS O FILICÍNEAS
Caracteres. — Los helechos viven generalmente en lugares sombríos y húmedos, por lo cual los
bosques, sobre todo los tropicales, constituyen su habitación predilecta; allí es donde son más
abundantes y alcanzan mayor tamaño. Los helechos arborescentes (alsophila) tienen una altura que
puede pasar de 10 metros.
El tallo es frecuentemente subterráneo y
tiene la forma de un rizoma, pero a veces
los hay erguidos como los precitados
heléchos arborescentes que terminan por un
penacho de grandes hojas, al estilo de las
palmeras. La periferia del cilindro presenta
una red de haces liberoleñosos unidos entre
sí, a expensas de los cuales se forman los
hacecillos que recorren las hojas. Los vasos
leñosos son escaleriformes.](https://image.slidesharecdn.com/botanicatodo-140511175115-phpapp01/85/Botanica-todo-106-320.jpg)
![107
Las frondas suelen ser de gran tamaño, muy divididas y delgadas, de modo que su parénquima, rico
en clorofila, puede obtener la energía necesaria para su actividad vital, a pesar de la escasez de los
rayos luminosos que reciben. Cuando la hoja es joven se encuentra arrollada en espiral', esto
representa una buena protección, pues le permite atravesar el suelo o las ramas de otras plantas sin
sufrir heridas que pudieran perjudicarle.
Tipo de estudio: el polipodio (Polypodium vulgare).
- Aparato vegetativo del helecho. El polipodio es una planta
vigorosa que abunda en los sitios húmedos y sombreados.
Frondas.- La porción aérea de la planta consta únicamente de grandes
frondas de verde intenso. Esas frondas van recortadas en numerosos
folíolos y éstos en foliolillos, que dan a cada folíolo la forma de peine
doble. Las nuevas hojas brotan por manojos, y salen enroscadas en
forma de cayados escamosos y velludos. A medida que van creciendo
las hojas se desenroscan y e] limbo se extiende y ensancha.
Tallos.- Si cavamos al pie de las hojas, notaremos que se desprenden de
un tallo subterráneo, cubierto de escamas rojizas: ese tallo es, pues, un rizoma. Se va ramificando
lateralmente y lleva en su porción inferior numerosas raíces adventicias endebles y negruzcas.
En esas raíces, así como en las hojas, podemos observar la existencia de tubitos o vasos conductores
de la savia.
El aparato vegetativo del helecho consta de raíces, tallo y hojas, con vasos conductores de la savia.
Aplicaciones.—Algunos heléchos como el polipodio y el culandrillo se cultivan mucho como
plantas de adorno en los parques y en los jardines. Otras especies exóticas y más delicadas se
cultivan en invernáculos, pues requieren poca luz, un ambiente turbio y húmedo y un suelo flojo y
rico en humus. Además, varios son empleados en farmacia por sus propiedades tónicas,
estimulantes y antihelmínticas. El rizoma del helecho se emplea como el remedio más eficaz contra
la tenia solitaria (Tenia solium). La escolopendra, o lengua de ciervo, es empleada como
astringente. Algunas variedades son muy afamadas; como hemostáticos, en Extremo Oriente.
Reproducción.
Los heléchos (y demás teridofitas) tienen
una reproducción alternante; es decir,
presentan dos ciclos, uno asexual y otro
sexual, que se suceden regularmente; al
primero se le llama esporofita y al segundo
gametofito.
Formación de las esporas.- Esporangio.-
Durante la primavera se puede observar en
la cara inferior de las hojas manchas
pardas, llamadas soros (A), protegidas a
menudo por un repliegue membranoso
que recibe el nombre de indusio (B). Cada
soro está constituido por un conjunto de
pequeños órganos (esporangios) que
tienen la forma de un pelo abultado en un
extremo (C) donde se hallan ciertas células
que forman las esporas, por un](https://image.slidesharecdn.com/botanicatodo-140511175115-phpapp01/85/Botanica-todo-107-320.jpg)




El documento presenta una introducción a la botánica como ciencia. Explica que la botánica estudia los vegetales y se divide en botánica general, especial y aplicada. La botánica general incluye la citología, histología, morfología, anatomía, fisiología, ecología y genética. La botánica especial se enfoca en la taxonomía, fitografía y fitogeografía. Finalmente, la botánica aplicada estudia la utilidad de las plantas en la alimentación, industria y medicina.










![Botanica1[1]](https://cdn.slidesharecdn.com/ss_thumbnails/botanica11-100818112519-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)


























![El cultivo del_repollo[1]](https://cdn.slidesharecdn.com/ss_thumbnails/elcultivodelrepollo1-151019001508-lva1-app6891-thumbnail.jpg?width=640&height=640&fit=bounds)








![59537682 manual-tecnico-de-hidroponia-popular[1]](https://cdn.slidesharecdn.com/ss_thumbnails/59537682-manual-tecnico-de-hidroponia-popular1-110914021538-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)








































