Descargado 448 veces









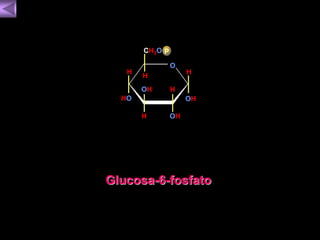


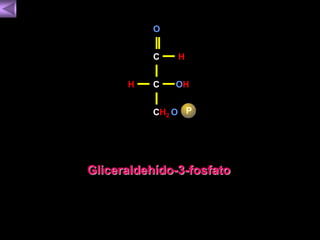
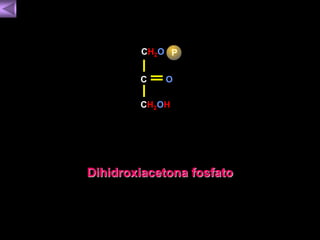



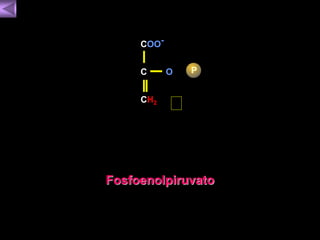

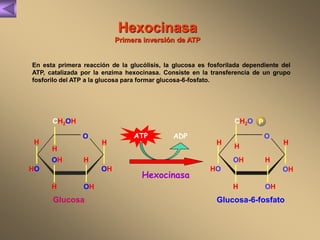







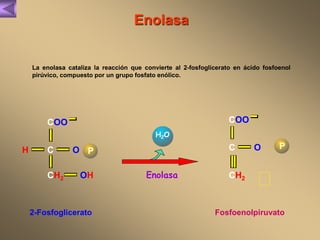











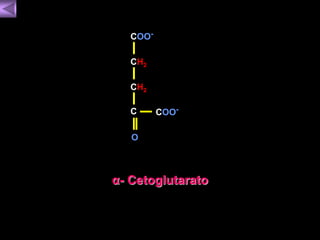








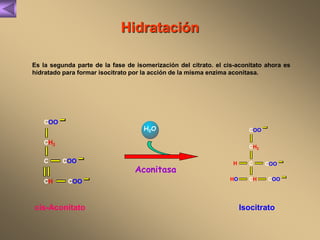








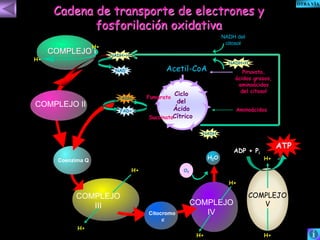





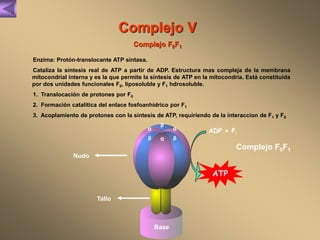









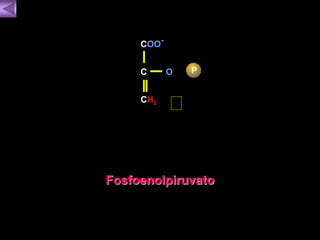



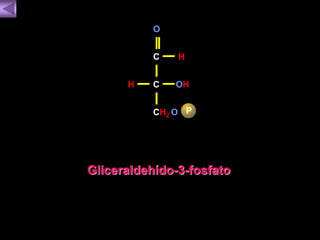
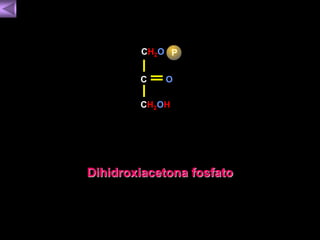

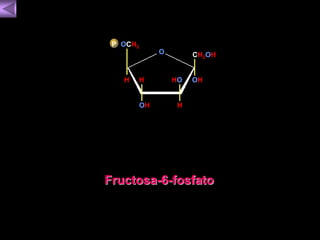
















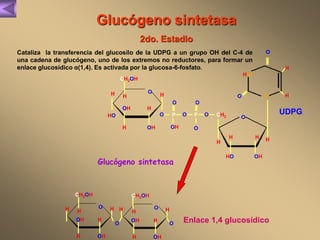
































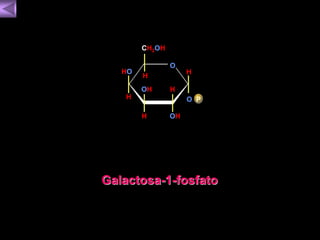



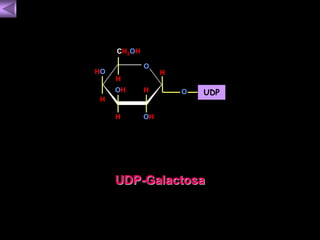



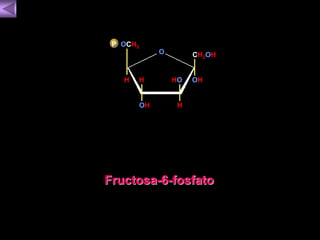
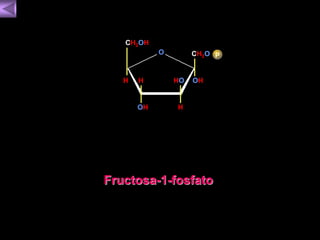





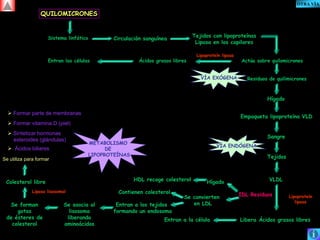























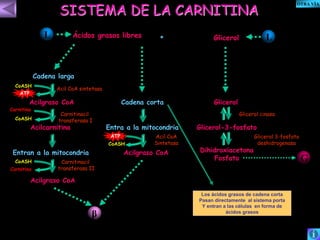



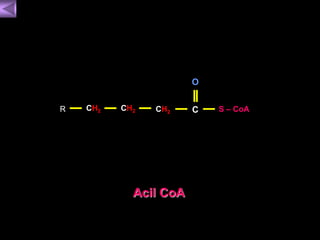








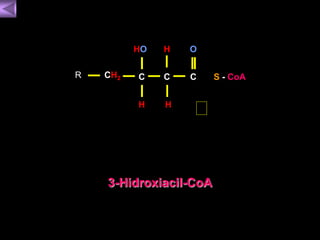




























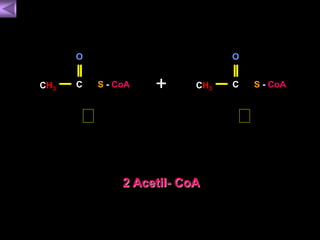






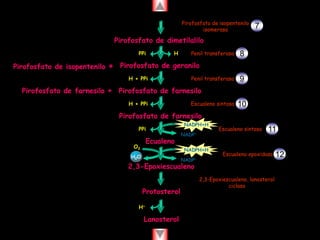
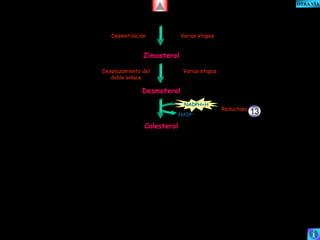








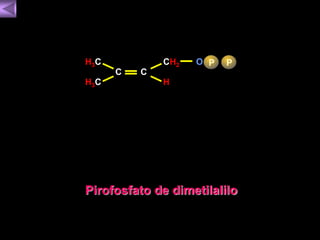




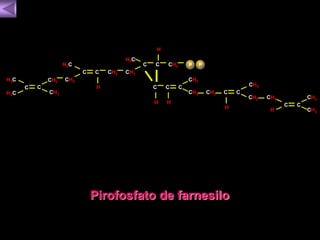





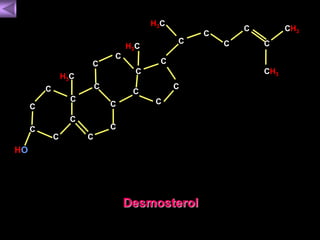









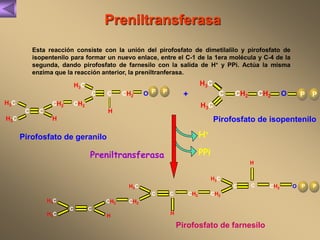



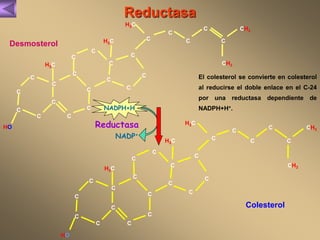




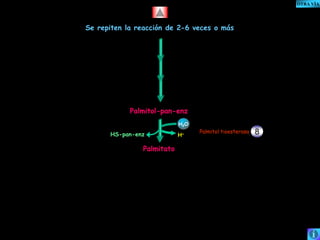













































El documento presenta un diagrama del ciclo del ácido cítrico y las rutas metabólicas asociadas. Muestra las 10 reacciones que convierten la glucosa en piruvato a través de la glucólisis, así como la entrada de piruvato, ácidos grasos y aminoácidos al ciclo del ácido cítrico. Finalmente, describe la oxidación del NADH a través de la cadena de transporte de electrones y la fosforilación oxidativa para generar ATP.















































