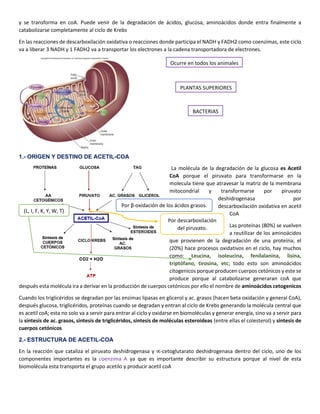
1. El ciclo de Krebs es una ruta metabólica central que participa en procesos catabólicos y anabólicos en la mitocondria. Catabólicamente oxida moléculas como carbohidratos, lípidos y proteínas para producir energía en forma de ATP, y anabólicamente genera precursores para la síntesis de nuevas moléculas.
2. El ciclo consta de 8 reacciones que degradan completamente la molécula de acetil-CoA derivada del metabolismo de glucosa, lípidos y