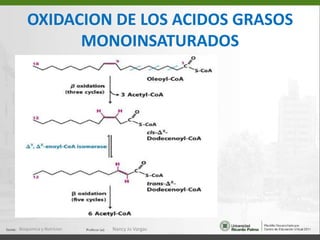
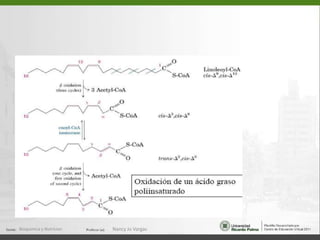
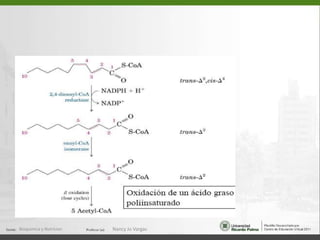
Este documento describe los procesos de beta-oxidación de ácidos grasos. Explica que los ácidos grasos son oxidados en la mitocondria para producir acetil-CoA a través de cuatro reacciones repetidas: oxidación, hidrólisis, oxidación y tiólisis. También cubre la oxidación de ácidos grasos insaturados y de cadena impar/par, así como la formación y uso de cuerpos cetónicos.